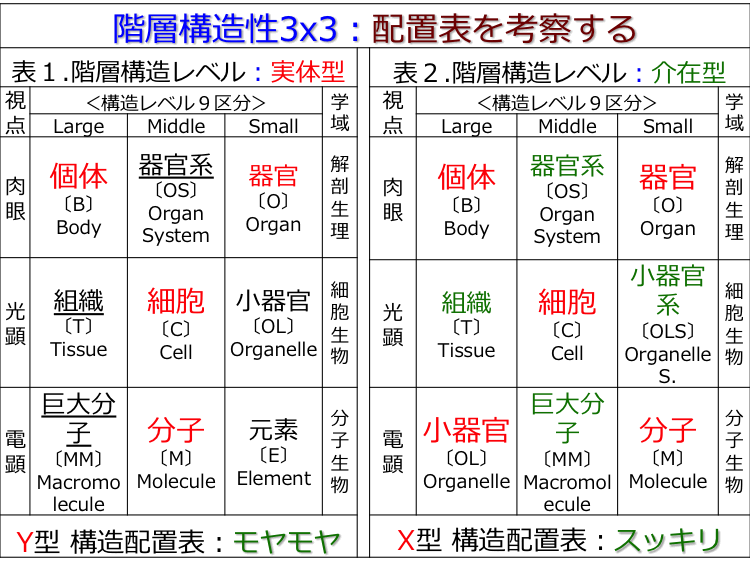
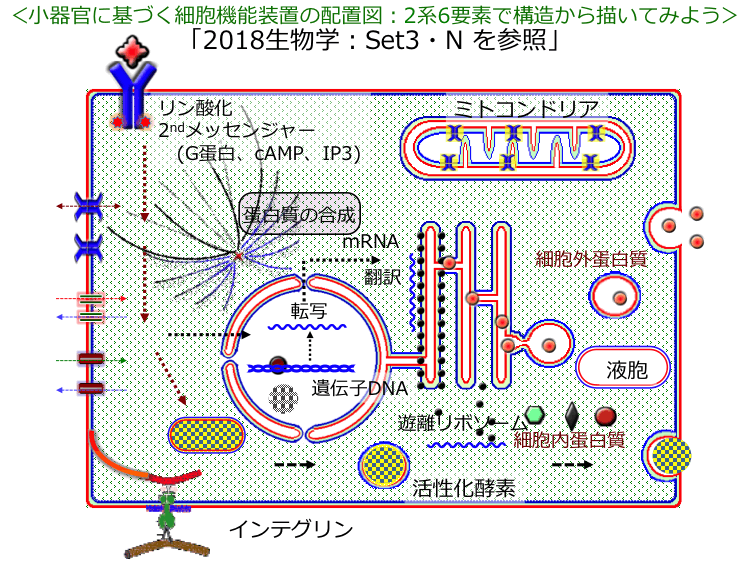
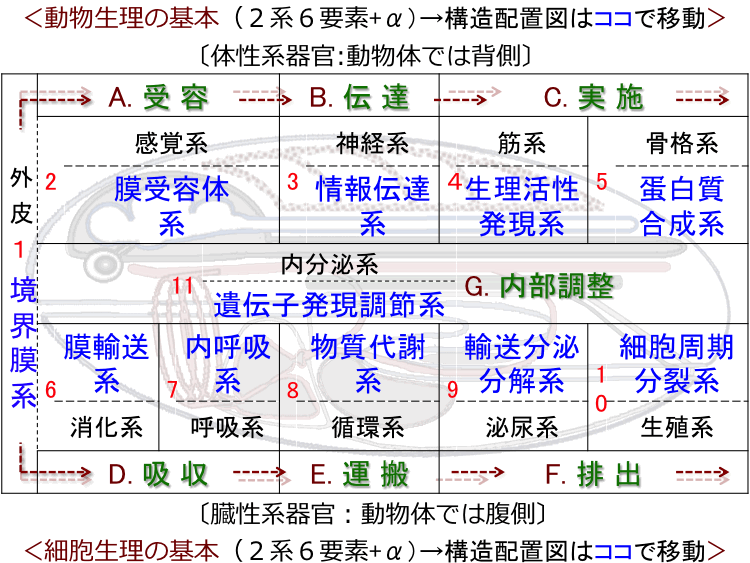
�� �� �M���̍\���́u�S5��, �S25��, �S109��, �S1317����, �S1595�Łv����
�@�i�ߍ��ڂ̕��������͂��̃e�L�X�g�̂��ߕt�L�������́B�����ɂ͂Ȃ���j�@ |
| �M���̖ڎ� |
�p���̖ڎ� |
| �\�� |
�זE�̕��q�����w |
Title |
Molecular Biology of the Cell |
| ��T�� |
�זE�Ƃ�
�i�R�� 8�� 136���� 196�Łj |
Part I |
Introduction to the Cell |
| �P�� |
�זE�ƃQ�m��
�i�\���F3�� 42���ځj
|
Ch.1 |
Cells and Genomes |
| �P��
�@�P�� |
1-1. �n����̍זE�����L��������i10���ځj
- ���ׂĂ̍זE�����w�����iDNA�j�Ɉ�`�������[���Ă��� p.2
- ���ׂĂ̍זE�́C���^��p�����d�������ň�`�������� p.3
- ���ׂĂ̍זE�͈�`���̋L����Ă���Ƃ����]�ʂ��ē������������ԑ́iRNA�j����� p.4
- ���ׂĂ̍זE�̓^���p�N����G�}�Ƃ��ėp���� p.5
- RNA ���^���p�N���ɖ|����@�͂��ׂĂ̍זE�œ��� p.6
- 1 �̃^���p�N���ɑΉ�������`���̈����`�q�Ƃ�� p.7
- �����͎��R�G�l���M�[��K�v�Ƃ��� p.8
- ���ׂĂ̍זE�͓�����{�\���P�ʕ��������������w�H��ł��� p.8
- ���ׂĂ̍זE�͍זE���ɕ�܂�Ă���C�h�{�f�ƘV�p���͂����ʉ߂���K�v������ p.9
- �זE�́C500 �ȉ��̈�`�q�Ő����Ă����� p.10
- �܂Ƃ� p.11
���擪�s���E�I��\�ցA�͐߂̖ڎ��� �A�\�Q����
|
| �P��
�@�Q�� |
1-2. �Q�m���̑��l���Ɛ����̌n�����i14���ځj�@
�@
- �זE�������܂��܂Ȏ��R�G�l���M�[�� p.12
- �זE�ɂ͒��f�Ɠ�_���Y�f���Œ肷����̂�����C�ق��̍זE������𗘗p���� p.13
- �����w�I�ȑ��l���́C���j�זE�ōł������� p.14
- �����̌n�����́C�ہC�ÍہC�^�j������3 �n�����Ȃ� p.15
- ��`�q�ɂ́C���₭�i��������̂ƁC�悭�ۑ��������̂����� p.16
- �ۂƌÍۂ�1000 �` 6000 �̈�`�q������ p.17
- �V������`�q�́C�����̈�`�q�������� p.18
- ��`�q�d���ɂ��C�זE�̒��ɗމ���`�q�̃t�@�~���[���ł��� p.19
- �������ł����R�E�ł��C�����Ԃň�`�q�̎n�����N���� p.21
- ���ɂ��C����ł̈�`���̐����������N���� p.22
- ��`�q�̋@�\�͉���z�炩�Ȃ萄���ł��� p.22
- 200 �ȏ�̈�`�q�t�@�~���[���C�n������3 �啪�}�̂�����ɂ����݂��� p.23
- �ψق���`�q�̋@�\�𖾂炩�ɂ��� p.23
- ���q�����w�҂͑咰�ۂ��W���I�Ɍ������Ă��� p.24
- �܂Ƃ� p.26
���擪�s���E�I��\�ցA�͐߂̖ڎ��� �A�\�Q����
|
| �P��
�@�R�� |
1-3. �^�j�����̈�`����i18���ځj
- �^�j�זE�̋N���͂����炭�ߐH�҂��낤 p.26
- ���݂̐^�j�זE�͋����ɂ���Đi������ p.27
- �^�j�����͋N���̓�����������Q�m�������� p.30
- �^�j�Q�m���͑傫�� p.30
- �^�j�Q�m���ɂ͒���DNA ����ʂɊ܂܂�Ă��� p.31
- �Q�m�����C���זE�����̔����v���O���������߂� p.31
- �^�j�����̑����͒P�זE�̌��������ł��� p.32
- �y��͍ŏ��̃��f���^�j�����ł��� p.33
- 1 �̐����̑S��`�q�̔������x���������Ɍv���ł��� p.34
- �זE�̈Ӗ���m��ɂ͐��w�C�v�Z�@�C��ʓI��K�v�ł��� p.35
- �V���C�k�i�Y�i��30 ����̂Ȃ����烂�f���A���Ƃ��đI�ꂽ p.36
- �����זE�E�̑�\�͐����C�n�G�C�}�E�X�C�q�g p.36
- �V���E�W���E�o�G�̌����͐Ғœ����̔����ׂ錮�ƂȂ� p.37
- �Ғœ����̃Q�m���͏d���̌J��Ԃ��̏��Y�ł��� p.38
- ��`�I�]��͈�`�w�҂ɂƂ��ē�肾���C�����i���ɂ͍D�܂��� p.39
- �}�E�X�͚M���ނ̃��f�������ł��� p.39
- �q�g�͎����̓Ɠ��ȓ_��ł��� p.40
- �ו����݂�C�����݂͂�Ȉ���Ă��� p.41
- �܂Ƃ� p.42
�͖���� p.42
���@�� p.44
���擪�s���E�I��\�ցA�͐߂̖ڎ��� �A�\�Q���� |
| �Q�� |
�זE�̉��w�Ɛ�����
�i�\���F3�� 43���ځj |
Ch.2 |
Cell Chemistry and Biosynthesis |
| �Q��
�@�P�� |
2-1. �זE�̉��w�g���i16���ځj�@
�@
- �זE�͏��Ȃ���ނ̌��q����ł��Ă��� p.45
- �ŊO�k�d�q�����q�Ԃ̑��ݍ�p�����߂� p.46
- ���L�����͓d�q�̋��L�ɂ���Ăł��� p.48
- ���L�����ɂ͂������̎�ނ����� p.50
- ���q�͌��܂������a�������̂悤�ɂӂ�܂��@p.51
- ���͍זE���ōł��L�x�ȕ����ł���@p.51
- �ɐ����q�ɂ͎_�≖��ƂȂ���̂�����@p.52
- �זE���ł�4��ނ̔L���͂����q�����т��Ă���@p.53
- �זE�͒Y�f����������ł��Ă���@p.54
- �זE���ɂ͂����ȏ��L�@���q�Q��4���� p.55
- ���͍זE�̃G�l���M�[���ł���C�����̍\���P�ʂł����� p.55
- ���b�_�̓G�l���M�[�������C�זE���̐����ł����� p.58
- �A�~�m�_�̓^���p�N���̍\���P�ʂł��� p.59
- �k�N���I�`�h��DNA��RNA�̍\���P�ʂł��� p.61
- �זE�̉��w�����́C���ڂɒl��������������啪�q���������Ă���@p.62
- �L�����ɂ���ċ��啪�q�̐��m�Ȍ`�Ƃق��̕��q�Ƃ̌��������܂�@p.63
- �܂Ƃ߁@p.65
���擪�s���E�I��\�ցA�͐߂̖ڎ��� �A�\�Q����
|
| 2��
�@�Q�� |
2-2. �זE�̍s���G�}�����ƃG�l���M�[���p�i17���ځj�@ p.65�@
- �זE�̑�ӂ͍y�f�������ǂ��Ă���@p.66
- �����̒����́C�זE���M�G�l���M�[����o���邱�ƂŐ��܂��@p.66
- �����������͑��z���𗘗p���ėL�@���q����������@p.68
- �זE�͗L�@���q�̎_���ɂ���ăG�l���M�[��@p.70
- �_���ƊҌ��ɂ͓d�q�̈ړ����������@p.71
- �y�f�͉��w�����̏�ǂ�Ⴍ����@p.72
- �y�f����Əo����@ �� ���q�^���̖җ�ȑ��� p.74
- �������N���邩�ǂ����͎��R�G�l���M�[�̕ω��Ō��܂�@p.75
- �������̔Z�x�ɂ���Ď��R�G�l���M�[�ω��Ɣ����̕������ς��@p.76
- �A�����ċN���锽���ɂ��Ă�DG8�����Z�ł���@p.77
- �����^���̕��q�͐������ɕs���ł���@p.78
- �����^���̂̓G�l���M�[�I�ɋN����₷�������Ƌ������Đ�������@p.79
- ATP �͍ł��悭�g���銈���^���̕��q�ł���@p.80
- ATP �ɒ~����ꂽ�G�l���M�[��2 �̕��q�̌����ɂ悭�g����@p.81
- NADH ��NADPH �͏d�v�ȓd�q�^���̂ł���@p.82
- �זE���ɂ͂ق��ɂ������̊����^���̕��q������@p.83
- ���̍����q�̍�����ATP �̉��������Ői�ށ@p.84
- �܂Ƃ߁@p.87
���擪�s���E�I��\�ցA�͐߂̖ڎ��� �A�\�Q����
|
| 2��
�@3�� |
2-3. �H������̃G�l���M�[�l���i10���ځj�@ p.88�@
- ��ATP �����̒��S�I�o�H�ł���@p.88
- ���y�ł͎_�f�Ȃ���ATP �������@p.89
- ������ƁC�_���ƃG�l���M�[�������y�f�����������邵���݂��킩��@p.91
- �����͐H�����q�����̒����ɂɒ~����@p.91
- �قƂ�ǂ̓����זE�́C�H���ƐH���̊ԂɎ��b�_����G�l���M�[�����o���@p.95
- ���Ǝ��b�͂ǂ�����~�g�R���h���A�ŃA�Z�`��CoA �ɕ��������@p.96
- �N�G���_��H�̓A�Z�`�����CO2 �Ɏ_������NADH �����@p.97
- �זE�̂قƂ�ǂ�ATP �͓d�q�`�B�ɂ���č��������@p.100
- �A�~�m�_�ƃk�N���I�`�h�͒��f�z�n�̈ꕔ�ł���@p.100
- ��ӂ͑g�D������C���߂���Ă���@p.101
- �܂Ƃ߁@p.103
�͖����@p.103
���@�� p.124
���擪�s���E�I��\�ցA�͐߂̖ڎ��� �A�\�Q����
|
| �R�� |
�^���p�N��
�i�\���F2�� 51���ځj
|
Ch.3 |
Proteins |
| 3��
�@�P�� |
3-1. �^���p�N���̌`�ƍ\���i18���ځj�@ p.125�@
- �^���p�N���q�̌`�̓A�~�m�_�z��ɂ���Č��܂�@p.125
- �^���p�N���̓G�l���M�[�Œ�̃R���z���[�V�����ɐ܂肽���܂��@p.130
- ���ʂ���܂肽���݃p�^�[���Cα�w���b�N�X��β�V�[�g�@p.131
- �^���p�N���q�͂����菬���ȃ^���p�N�h���C���̑g�ݍ��킹�ō����@p.135
- ���_�I�ɉ\�ȃ|���y�v�`�h���̂Ȃ��ōזE�ɂƂ��ėL�p�Ȃ��͔̂�r�I���Ȃ��@p.136
- �^���p�N���͑����̃t�@�~���[�ɕ��ނł���@p.137
- �z���������ŋ߉����ł���@p.139
- ���܂��܂ȃ^���p�N���̕��i�ƂȂ�^���p�N�h���C���C���W���[���@p.140
- �����̃^���p�N���ɑɂȂ����h���C��������@p.141
- �q�g�Q�m�����w�߂��镡�G�ȃ^���p�N�Q�̑����͂܂��𖾂���Ă��Ȃ��@p.142
- ��^�^���p�N���q�̑����͕����̃|���v�`�h������Ȃ�@p.142
- �����点���̑@�ۂ����^���p�N��������@p.143
- �̂т��@�ۏ�\�����Ƃ�^���p�N������������@p.145
- �����̃^���p�N���ŁC���m�ȍ\���̂Ȃ��|���y�v�`�h�������������قǑ����@p.146
- �זE�O�̃^���p�N���͋��L�����ɂ��ˋ��ň��艻����@p.147
- �^���p�N�����T�u���j�b�g�Ƃ��ďW�����傫�ȍ\���̂ɂȂ�@p.148
- �זE���̍\���̂̑����͎��ȏW���ł���@p.149
- ���G�Ȑ��̍\���͏W�����q�̏�������Č`������邱�Ƃ������@p.151
- �܂Ƃ߁@p.152
���擪�s���E�I��\�ցA�͐߂̖ڎ��� �A�\�Q����
|
| 3��
�@�Q�� |
3-2.�@�^���p�N���̋@�\�i33���ځj�@ p.152
- ���̕��q�ƌ������Ȃ��^���p�N���͂Ȃ��@p.153
- �^���p�N���̕��q�\�ʂ̃R���z���[�V���������̕��q�̉��w�I��
- �������߂�@p.154
- �����̃^���p�N���ǂ����̔z���r�ɂ��C�d�v�ȃ��K���h�������ʂ������яオ��@p.155
- �^���p�N���Ԃ̌����ɂ͂��܂��܂ȐڐG�l��������@p.156
- �R�̂̌������ʂ͂Ƃ�킯�Z�ʐ��ɕx�ށ@p.156
- ���t�萔�͌����̋����̎ړx�ł���@p.157
- �y�f�͋��͂ł���߂ē��ِ��̍����G�}�ł���@p.158
- ��Ƃ̌������y�f�̐G�}�����̑�1 �i�K�ł���@p.159
- �y�f�͑J�ڏ�Ԃ�I��I�Ɉ��艻���Ĕ������x���グ��@p.160
- �y�f�͎_�G�}�����Ɖ���G�}�������ɍs����@p.160
- ���]�`�[�����ɂ��Ă݂�y�f��p�̂����݁@p.161
- �^���p�N���ɋ����������Ă��鏬���q������̋@�\��t������@p.166
- �����̐G�}���ʂ����y�f�͊�q�g���l���ʼn^�ԁ@p.167
- �����y�f�n�̓����ōזE�̑�ӑ��x�������@p.168
- �זE���y�f�̐G�}�����߂��Ă���@p.169
- �A���X�e���b�N�y�f�ɂ͑��݂ɍ�p������2 �ȏ�̌������ʂ�����@p.171
- �������ʂ��������Ă���2 ��ނ̃��K���h�́C�������̌����ɉe���������@p.171
- �Ώ̓I�ȑg�ݗ��Ẵ^���p�N���͋����I�ɃA���X�e���b�N�]�ڂ���@p.172
- �A�X�p���M���_�J���o���C����]�ڍy�f�̃A���X�e���b�N�]�ڂ͌��q���x���ʼn𖾂���Ă���@p.173
- �^���p�N���̕ω��̑����̓^���p�N���̃����_���ŋN����@p.175
- �^�j�זE�ɂ͑���̃^���p�N�L�i�[�[��^���p�N�z�X�t�@�^�[�[�����݂���@p.176
- Cdk ��Src �^���p�N�L�i�[�[�̒��߂���C�^���p�N�����}�C�N���`�b�v�Ƃ��ċ@�\���邵���݂��킩��@p.177
- GTP ��������������������^���p�N���́C���߈��q�Ƃ��čזE���ɍL�����݂��Ă���@p.178_
- GTP �����^���p�N��GTP ��GDP �̂ǂ��炪�������Ă��邩�߃^���p�N�͌�����߂�@p.179
- �^���p�N���̑傫�ȓ����͏����ȓ������琶�ݏo�����@p.179
- ���[�^�[�^���p�N�͍זE���ő傫�ȓ����ݏo���@p.181
- �������A���͖̂���ʂ������q�̈ړ��ɃG�l���M�[�𗘗p����@p.182
- �^���p�N������^�̕����̂�����ă^���p�N���u�Ƃ��ċ@�\���邱�Ƃ��悭����@p.184
- �^���p�N���u�͌����ł��镔�i���g����`���������悭���p����@p.184
- �^���p�N���u������̏ꏊ�ւ̔z�u���������ɂȂ���@p.185
- �^���p�N���̑����͕������ʂ̋��L�����C���ɂ�蒲�߂����@p.186
- ���G�ȃ^���p�N���ݍ�p�Ԃ��זE�̋@�\�̊�b�ƂȂ�@p.187
- �܂Ƃ߁@p.190
�͖����@p.191
���@���@p.193
���擪�s���E�I��\�ցA�͐߂̖ڎ��� �A�\�Q����
|
| ��U�� |
��`�̊�{ |
Part II |
Basic Genetic Mechanisms |
| �S�� |
DNA�C���F�́C�Q�m��
�i�\���F4�� 45���ځj |
Ch.4 |
DNA and Chromosomes |
| 4��
�@�P�� |
4-1. DNA�̍\���Ƌ@�\�i3���ځj�@ p.197�@
- DNA ���q��2 �{�̑���I�ȃk�N���I�`�h���łł��Ă���@p.197
- DNA �̍\�������`�̂����݂����܂�@p.199
- �^�j������DNA �͊j���ɋǍ݂���@p.200
- �܂Ƃ߁@p.201
���擪�s���E�I��\�ցA�͐߂̖ڎ��� �A�\�Q����
|
| 4��
�@�Q�� |
4-2. ���F��DNA�Ƃ��̃N���}�`���@�ۂւ̋l�ߍ����i11���ځj�@ p.202
- �^�j������DNA �͐��F�̂̃Z�b�g�Ɏ��߂��Ă���@p.202
- ���F�̂ɂ͒�����Ȃ�����`�q���܂܂�Ă���@p.204
- �q�g�Q�m���̉���z��C�q�g�ł̈�`�q�̔z�u���킩��@p.205
- �Q�m���̔�r����C�i���ɂ����ĕۑ�����Ă�������z�킩��@p.207
- ���F�͍̂זE�����̒i�K�ɂ���ĈقȂ��Ԃ��Ƃ��đ��݂���@p.208
- ����̐��F�̂��`�����Ă���DNA ���ɂ́C�K��1 �̃Z���g�����A�C2 �̃e�����A�ƕ����N�_������@p.209
- ���F�̓��ł�DNA ���q�͍��x�ɋÏk���Ă���@p.210
- �k�N���I�\�[���͐^�j�����̐��F�̍\���̊�{�P�ʂł���@p.211
- �k�N���I�\�[���E�R�A���q�̍\������DNA ���l�ߍ��ނ����݂��킩��@p.212
- �k�N���I�\�[���\���͓��I�ŁCATP �ˑ��N���}�`���č\�������̂ɂ���č\����ω������@p.215
- �k�N���I�\�[�����܂Ƃ܂��ċÏk���N���}�`���@�ۂ��ł���@p.216
- �܂Ƃ߁@p.218
���擪�s���E�I��\�ցA�͐߂̖ڎ��� �A�\�Q����
|
| 4��
�@�R�� |
4-3. �N���}�`���\���̒����i10���ځj�@ p.219
- �����ɂ�����N���}�`���\���̓�@p.220
- �w�e���N���}�`���͍����\�����Ƃ��Ă���C��`�q�������ُ�ɗ}���Ă���@p.220
- �R�A�E�q�X�g���͂��낢��ȏꏊ�ŋ��L�����C������@p.222
- �����̕ώ�q�X�g�������ٓI�ʒu�ɑ}������āC�N���}�`���͂���ɑ��l�ɂȂ�@p.224
- ���L�����C���ƕώ�q�X�g���̋����ŁC�����̋@�\�����߂�“�q�X�g���R�[�h”�����@p.224
- �R�[�h�ǂݎ��^���p�N�ƃR�[�h�������݃^���p�N�̕����̂����F�̏�ɋN�����N���}�`���̏C���������܂ōL�߂�@p.226
- ��ǔz��DNA ���אڂ���N���}�`���̈�ɓǂݏ��������̂��L����̂�W����@p.227
- �Z���g�����A�̃N���}�`������C�ώ�q�X�g��������̍\������邵���݂����炩�ɂȂ�@p.228
- �N���}�`���\���̒��ڌp�������肤��@p.230
- �^�j�זE�̐��F�̂̋@�\�ɌŗL�̐����̓N���}�`���\���ɗR������@p.231
- �܂Ƃ߁@p.233
���擪�s���E�I��\�ցA�͐߂̖ڎ��� �A�\�Q����
|
| 4��
�@�S�� |
4-4. ���F�̂̑S�̍\���i7���ځj�@ p.233
- ���F�̂͐܂肽���܂�āC�N���}�`���̑傫�ȃ��[�v�����@p.234
- �������F�̂̓N���}�`���\���̎��o���ɓ��ɗL�p�ł���@p.236
- ���낢��Ȍ`�Ԃ̃w�e���N���}�`��������@p.238
- �N���}�`�����[�v�͒��̈�`�q����������Ƃ��Ïk�x���Ⴍ�Ȃ�@p.239
- ��`�q������ς��邽�߂ɁC�N���}�`���͊j�̒��œ���̈ʒu�Ɉړ����邱�Ƃ�����@p.239
- ��^���q�̖Ԗڍ\�����j�����w�I�ɓ����̂���̈�Q�ɕ����Ă���@p.241
- �L�����̐��F�͍̂ł��Ïk�����N���}�`���łł��Ă���@p.243
- �܂Ƃ߁@p.245
���擪�s���E�I��\�ցA�͐߂̖ڎ��� �A�\�Q����
|
| 4��
�@�T�� |
4-5. �Q�m���i���̂������i14���ځj�@ p.246
- �Q�m���̕ω���DNA �̕����ƈێ��̋@�\�̎��s�ɂ�萶����@p.246
- ��̐����Ԃ̃Q�m������z��̈Ⴂ�́C����炪�i���̉ߒ��ŕ��Ă���̎��Ԃ̒����ɔ�Ⴗ��@p.247
- DNA ����z���r����ɂ����n��������C�����鐶���̊W�����ǂ��@p.248
- �q�g�ƃ}�E�X�̐��F�̂��r����ƁC�Q�m���\���̕���̂����݂��킩��@p.249
- �Ғœ����̃Q�m���̑傫���́C���̐������DNA �l���Ƒr���̑��ΓI���x�f����@p.251
- �����c��̃Q�m����DNA ����z����Č��ł���@p.251
- ����Ԃ�DNA ����z���r�ɂ��C�@�\���m�����d�v��DNA ����z���ł���@p.252
- �q�g�Œ����ۑ�����Ă����̂ɋߔN�}���ȕω�������������z��́C�q�g�̐i����̂���߂ďd�v�Ȓi�K�̌����ɂ���@p.253
- ��`�q�̏d���͐i���ɂ����ĐV�������̂����o�����ƂȂ�@p.253
- �d����`�q�̑��l���@p.254
- �O���r����`�q�t�@�~���[�̐i���́CDNA �d���������̐i���Ɋ�^���邵���݂������Ă���@p.256
- �G�L�\���Ԃ̑g�݊����ɂ���ĐV������`�q�������\��������@p.257
- �����ψق͏W�c�ɍL�܂�C���̌̐��ɉ������m���ŌŒ肳���@p.257
- �q�g�ɂ�����ψق̉𖾂ł��낢��Ȃ��Ƃ��킩��@p.258
- �܂Ƃ߁@p.260
�͖����@p.260
���@���@p.262
���擪�s���E�I��\�ցA�͐߂̖ڎ��� �A�\�Q����
|
�T�� |
DNA�̕����C�C���C�g����
�i�\���F7�� 56���ځj |
Ch.5 |
DNA Replication, Repair, and Recombination |
| 5��
�@�P�� |
5-1. DNA����z��̈ێ��i3���ځj�@ p.263
- �ψق��N����m���͋ɒ[�ɒႢ�@p.263
- �����̂ɂƂ��ĕψٗ��͏������Ȃ���Ȃ�Ȃ��@p.265
- �܂Ƃ߁@p.265
���擪�s���E�I��\�ցA�͐߂̖ڎ��� �A�\�Q����
|
| 5��
�@�Q�� |
5-2. DNA�����@�\�i11���ځj�@ p.266
- ����Ό`�����CDNA ������DNA �C���̊�{�ɂȂ��Ă���@p.266
- DNA �����t�H�[�N�͔�Ώ̂ł���@p.266
- DNA �����̍��������x�ɂ͍Z���@�\���K�v�ł���@p.268
- DNA ������5’ → 3’ �����łȂ��ƌ����I�ȍZ���͂ł��Ȃ��@p.271
- ����̃k�N���I�`�h�d���y�f�����M���O���p�̒Z���v���C�}�[RNA ���q����������@p.272
- �����t�H�[�N�̑O����DNA�����J���̂����������̃^���p�N��������@p.273
- ����\���̂������ŁCDNA �|�������[�[���q��DNA ���痣�ꂸ�Ɉړ�����@p.273
- �����t�H�[�N�ɂ����X�̃^���p�N�����������ĕ������u���`�����Ă���@p.275
- �������u�����̂����������̌��́C�s�����̑I��I�C���n����菜���@p.276
- ��������DNA ������Ȃ��悤�CDNA �g�|�C�\�����[�[�������@p.278
- DNA �����͐^�j�����ł��ۂł���{�I�ɂقړ����ł���@p.280
- �܂Ƃ߁@p.281
���擪�s���E�I��\�ցA�͐߂̖ڎ��� �A�\�Q����
|
| 5��
�@3�� |
5-3. DNA�����̊J�n�Ɗ����i13���ځj�@ p.281
- DNA �����͕����N�_�Ŏn�܂�@p.281
- �ۂ̐��F�̂ɂ͒ʏ�DNA�����N�_��1��������Bp.282
- �^�j�����̐��F�̂ɂ͕����̕����N�_������Bp.282
- �^�j�����ł͍זE�����̈ꎞ�������Ɍ�����DNA �������N����Bp.284
- ������F�̂ɂ����Ă��C�قȂ�̈��S ���̈قȂ鎞���ɕ��������Bp.285
- �Ïk�x�̒Ⴂ�N���}�`���̈�`�q�͏����ɕ�������C���x�ɋÏk�����N���}�`���͌���ɕ��������Bp.285
- �P���Ȑ^�j�����ł���o��y��Œ��ׂ�ƁC����̉���z�����N�_�Ƃ��ē����Bp.286
- �����̃T�u���j�b�g����Ȃ�傫�ȕ����̂��^�j�����̕����N�_�Ɍ�������Bp.287
- �����J�n���w������M���ނ̉���z��͓��肪�ނ��������Bp.288
- �V�����k�N���I�\�[���������t�H�[�N�̂�����Ō`�������Bp.289
- �^�j�����̐��F�̕����@�\�̂������ŁC�q�X�g���̏C���p�^�[���͊m���Ɏp�����Bp.290
- �e�������[�[�����F�̖̂��[������Bp.292
- �e�����A�̒����͍זE�ɂ��C�܂������ɂ�蒲�߂���Ă���Bp.293
- �܂Ƃ߁Bp.294
���擪�s���E�I��\�ցA�͐߂̖ڎ��� �A�\�Q����
|
| 5��
�@�S�� |
5-4. DNA�C���i8���ځj 295
- DNA �C�����Ȃ���C�����I�ȑ����ɂ����DNA �̉���z��͋}���ɕω����邾�낤 p.296
- DNA ��d�点��͗e�ՂɏC���ł��� p.296
- DNA �����̏����ɂ͕����̓������� p.297
- DNA �C���Ɠ]�ʂ��������āC�זE�̍ł��d�v��DNA �������悭�C������ p.299
- DNA ����̉��w�I�����������̔�����e�Ղɂ��� p.300
- �ً}���ɂ͓����DNA �|�������[�[���g����DNA ���C������ p.302
- ��{���ؒf�͌����悭�C������� p.302
- DNA �̑����͍זE�����̐i�s��x�点�� p.303
- �܂Ƃ� p.304
���擪�s���E�I��\�ցA�͐߂̖ڎ��� �A�\�Q�������擪�s���E�I��\�ցA�͐߂̖ڎ��� �A�\�Q����
|
| 5��
�@�T�� |
5-5. �����g�����i11���ځj�@ p.304
- �����g�����͍זE���ł��܂��܂ȗp�r�Ɏg���� p.304
- �����g�����̓����́C������זE�ɋ��ʂ��Ă��� p.305
- DNA �̉���Ό`���������g������U������ p.305
- RecA �^���p�N�₻�̑����̂́C��{����DNA ���d�点������ɂ��鑊���ȗ̈�Ƒ������� p.307
- ����_�ړ��ɂ��C�w�e����d���̈悪�g�傷�邩�C�V���ɍ������ꂽDNA ����{���Ƃ��ėV������ p.308
- �����g�����ɂ����DNA �̓�{���ؒf�����Ȃ��C���ł��� p.308
- �זE�́CDNA �C���ւ̑����g�����̗��p��T�d�ɐ��䂵�Ă���p.310
- �����g�����̍ۂɂ́C�����Ƀz���f�C�\�����`�������p.311
- ���������g�����̓v���O�������ꂽ��{���ؒf����n�܂�p.312
- �����g�����͈�`�q�ϊ��̌����ƂȂ邱�Ƃ�����p.314
- ���܂�K�����Ȃ�DNA �̊Ԃŕs�K�Ȉ�`�I�g�������N����Ȃ��悤�C�s���Z���n������p.315
- �܂Ƃ� p.316
���擪�s���E�I��\�ցA�͐߂̖ڎ��� �A�\�Q����
|
| 5��
�@6�� |
5-6. �]�ڂƕۑ��^���ʓ��ٓI�g�����i10���ځj�@ p.316
- ������`���q�͓]�ڋ@�\�ɂ���Ăǂ��DNA �z��̒��ɂ����荞�߂�p.317
- DNA �^�g�����X�|�]���͐�\��@�\�ƕ����@�\�̗����ɂ��ړ�����p.317
- �]�ڋ@�\���g���ďh��זE�̐��F�̂ɓ��荞�ރE�C���X������p.319
- ���g���E�C���X�^���g���g�����X�|�]���̓��g���E�C���X�Ɏ��Ă��邪�C�^���p�N�O��������Ȃ�p.320
- �q�g�Q�m���̂��Ȃ�̕����́C�g���E�C���X�^���g���g�����X�|�]���ɐ�߂��Ă���p.321
- �ǂ�ȓ]�ڈ��q���������͐����ɂ���ĈقȂ�p.322
- �]�ڈ��q�̈ړ����N���������悻�̎����̓Q�m������z��ǂݎ���p.323
- �ۑ��^���ʓ��ٓI�g������DNA �̉t�I�ȍĕҐ����\�ɂ���p.323
- �ۑ��^���ʓ��ٓI�g�����̓o�N�e���I�t�@�[�Wl �Ŕ������ꂽp.324
- �ۑ��^���ʓ��ٓI�g�����𗘗p���Ĉ�`�q�̃X�C�b�`���I���E�I�t�ł���p.324
- �܂Ƃ�p.326
�͖����p.327
���@��p.328
���擪�s���E�I��\�ցA�͐߂̖ڎ��� �A�\�Q����
|
| �U�� |
�Q�m�����̓ǂݎ��
�FDNA����^���p�N����
�i�\���F3�� 55���ځj |
Ch.6 |
How Cells Read the Genome
�@: From DNA to Protein |
| 6��
�@�P�� |
6-1. DNA����RNA���i24���ځj�@ p.331
- DNA �̉���z��̈ꕔ��RNA �ɓ]�ʂ����p.332
- �]�ʂł́CDNA �̈���̍��ɑ���I��RNA �����@p.333
- �זE�ō����RNA �ɂ͐���ނ���@p.335
- DNA �ɂ́CRNA �|�������[�[���������n�߂�ꏊ�C�I����ꏊ���w������V�O�i��������@p.336
- �]�ʊJ�n�ƏI���������V�O�i���̉���z��͑��l�ł���@p.338
- �^�j�����ł̓]�ʊJ�n�ɂ͑����̃^���p�N�����K�v�ł���@p.339
- RNA�@�|�������[�[II �͓]�ʊ�{���q��K�v�Ƃ��� p.340_
- �|�������[�[II �́C�]�ʊ��������q�C��݈��q�C�N���}�`���C���^���p�N���K�v�Ƃ��� p.342
- �]�ʂ̐L���ɂ���āCDNA �ɂ͂˂���̒��͂������� p.343
- �^�j�����̓]�ʂ́CRNA �v���Z�V���O�Ɩ��ɘA�g���Đi�s���� p.345
- �^�j������mRNA �O��̂ɑ���ŏ��̏C�������́C�L���b�v�`���ł��� p.346
- RNA �X�v���C�V���O�ł͐V���ɓ]�ʂ��ꂽmRNA �O��̂���C���g��������������� p.347
- �X�v���C�V���O�̋N����ʒu�͉���z�w������ p.349
- RNA �X�v���C�V���O�̓X�v���C�\�\�[�����s�� p.349
- �X�v���C�\�\�[���͈�A�̕��G��RNA–RNA �ĕҐ����s�����߂�ATP ���������𗘗p���� p.351
- mRNA �O��̂̂ق��̓����₻�̍�����������K�ȃX�v���C�X���ʂ̑I���ɖ𗧂� p.352
- ������A���ɂ́C�ꕔ�̃C���g�����������������snRNP �Q������ p.353
- RNA �X�v���C�V���O�͋����قǏ_������� p.355
- �X�v���C�\�\�[�����G�}����RNA �X�v���C�V���O�́C���ȃX�v���C�V���O�@�\����i�������炵�� p.355
- �^�j����mRNA ��39 ���[��RNA �v���Z�V���O�y�f����� p.357
- �^�j�����̐��nmRNA �͊j����I��I�ɉ^�яo����� p.358
- �����̔�|��RNA ���j�ō�������C���H����� p.360
- �j���̂̓��{�\�[���������u�ł��� p.362
- �j�ɂ͂��܂��܂Ȋj���\�������݂��� p.363
- �܂Ƃ� p.366
���擪�s���E�I��\�ցA�͐߂̖ڎ��� �A�\�Q����
|
| 6��
�@�Q�� |
6-2. RNA����^���p�N�����i24���ځj�@ p.366
- mRNA �̉���z��̓k�N���I�`�h3 ���̑g�ݍ��킹�Ƃ��ēǂݎ���� p.367
- tRNA ���q��mRNA �̃R�h���ƃA�~�m�_�����т��� p.368
- tRNA �͊j����^�яo�����O�ɋ��L�����C������ p.369
- ���ٓI�y�f���A�~�m�_��Ή�����tRNA ���q�Ɍ���������@p.370
- tRNA �����y�f�ɂ��ҏW�Ő��x���ۂ���� 371
- �A�~�m�_�͐L�����̃|���y�v�`�h���̃J���{�L�V���[�ɕt�������@p.373
- RNA �̎w�߂̓��{�\�[���ʼn�ǂ���� p.373
- �L�����q�͖|��𑣐i���C���x�����߂� p.377
- ���{�\�[���̓��{�U�C���̈��ł��� p.378
- mRNA �̉���z�^���p�N�����̊J�n�_���w������ p.379
- �I�~�R�h�����|��I�����w������ p.381
- �^���p�N���̓|�����{�\�[���ō�������� p.381
- �W���I�Ȉ�`�Í��ɂ���������������^������ p.382
- ���j�����̃^���p�N�����j�Q�܂͍R�������Ƃ��Ė𗧂� p.383
- ���m�Ȗ|��ɂ͎��R�G�l���M�[�̏���K�v�ł��� 385
- ��ꂽDNA ���|��Ȃ��悤�ɁC�i���Ǘ��@�\������ p.385
- �ꕔ�̃^���p�N���̐܂肽���݂́C�����̍Œ��Ɏn�܂� p.387
- �قƂ�ǂ̃^���p�N���ł̓V���y�������܂肽���݂������� p.388
- �I�o�����a�����̈�̓^���p�N���̕i���Ǘ��ɕs���Ȗڈ�ƂȂ� p.390
- �v���e�A�\�[���́C�������ʂ��I�o���Ȃ��悤�v���e�A�[�[������߂����ł��� p.391
- �I���ȃ��r�L�`�������@�\���������ׂ��^���p�N���ɖڈ������ p.393
- �����̃^���p�N���́C���䂳�ꂽ�����ɂ���Ē��߂���Ă��� p.395
- �܂肽���ݕ���������^���p�N���͋ÏW�̂��`�����C�q�g�̏d��ȕa�C�̌����ɂȂ邱�Ƃ����� p.396
- DNA ����^���p�N�����ł���܂łɂ͐������̒i�K������ p.399
- �܂Ƃ� p.399
���擪�s���E�I��\�ցA�͐߂̖ڎ��� �A�\�Q����
|
| 6��
�@�R�� |
6-3. RNA���E�Ɛ����̋N���i7���ځj 400
- �����ɂ͕ۑ����ꂽ��K�v�ł��� p.401
- �|���k�N���I�`�h�͏���ۑ��ł��邤���ɉ��w������G�}�ł��� p.401
- RNA ���E�̑O�ɂ����炭��RNA ���E�����邾�낤 p.402
- ��{��RNA ���q�͂���߂ĕ��G�ȍ\�����Ƃ�� p.403
- ���ȕ������q�͎��R�I������ p.404
- �^���p�N�����͂ǂ̂悤�ɐi�������̂��낤 p.407
- �����̍זE�͈�`�����Ƃ���DNA �𗘗p���� p.408
- �܂Ƃ� p.408
�͖���� p.409
���@�� p.410
���擪�s���E�I��\�ցA�͐߂̖ڎ��� �A�\�Q����
|
| �V�� |
��`�q�����̒���
�i�\���F5�� 87���ځj |
Ch.7 |
Control of Gene Expression |
| 7��
�@�P�� |
7-1. ��`�q���߂̊T���i14���ځj�@ p.411
- ���זE�����ł͍זE�̌^������Ă�����DNA ������ p.411
- �זE�^�̈Ⴂ�͍�������^���p�N���̑g�ݍ��킹�̈Ⴂ�ɂ�� p.412
- �O������̃V�O�i�����זE�̈�`�q������ω������邱�Ƃ����� p.413
- ��`�q������DNA�CRNA�C�^���p�N���Ɛi�ތo�H�̂��낢��Ȓi�K�Œ��߂���� p.415
- �܂Ƃ� p.415
���擪�s���E�I��\�ցA�͐߂̖ڎ��� �A�\�Q����
|
| 7��
�@�Q�� |
7-2. ��`�q���߂ɓ����^���p�N����DNA�������`�[�t�i18���ځj�@ p.416
- ��`�q���߃^���p�N�́C�ۈ�`�w�̌����Ŕ������ꂽ p.416
- DNA �点��̊O�����^���p�N�����ǂ� p.416
- ��`�q�X�C�b�`�̊�{������DNA �̒Z������z��ł��� p.418
- ��`�q���߃^���p�N��DNA ����z���ǂݎ��\�����`�[�t������ p.418
- �w���b�N�X�|�^�[���|�w���b�N�X�iHTH�j���`�[�t�͍ł��P������ʓI��DNA �������`�[�t��1 �ł��� p.419
- �z���I�h���C���^���p�N�͓��ʂ�HTH �^���p�N�ł��� p.420
- DNA ����Zn�i�W���N�j�t�B���K�[�E���`�[�t�ɂ͐���ނ��� p.421
- β�V�[�g��DNA �����ʂ��� p.422
- �傫���a�Ə������a�ɓ��郋�[�v���g����DNA �����ʂ���^���p�N�������� �@p.423
- ���C�V���W�b�p�[�E���`�[�t��DNA �̌����ƃ^���p�N���̓�ʑ̌`���Ƃ�}��� �@p.423
- �w�e����ʑ̌`���ɂ���`�q���߃^���p�N�����ʂł��鉖��z��͈̔͂��L���� �@p.424
- �w���b�N�X�|���[�v�|�w���b�N�X�iHLH�j���`�[�t����ʑ̌`����DNA �����Ƃ�}��� �@p.425
- ��`�q���߃^���p�N�����ʂ��鉖��z������ׂė\�����邱�Ƃ͂܂��ł��Ȃ� �@p.426
- �Q���V�t�g�@�͉���z����ٓIDNA �����^���p�N��e�ՂɌ��o���� �@p.427
- DNA �A�t�B�j�e�B�[�N���}�g�O���t�B�[�ɂ��C����z����ٓIDNA �����^���p�N���ȒP�ɐ����ł��� p.428
- ��`�q���߃^���p�N�����ʂ���DNA ����z��͎����I�Ɍ���ł��� �@p.429
- �n�������w�I�t�b�g�v�����g�@�͔�r�Q�m����͂ɂ���Ē���DNA �肷�� p.431
- ���זE�̒��ň�`�q���߃^���p�N���������镔�ʂ̑����̓N���}�`���Ɖu���~�@�ł킩�� p.431
- �܂Ƃ� p.432
���擪�s���E�I��\�ցA�͐߂̖ڎ��� �A�\�Q����
|
7��
�@�R�� |
7-3. ��`�q�X�C�b�`�̓����������i19���ځj�@ p.432
- �ۂ̃g���v�g�t�@���E���v���b�T�[�͈�`�q���I���E�I�t���� p432.
- �P���ȃX�C�b�`�ł��� p.433
- ��`�q���I���ɂ���]�ʊ��������q 435
- �]�ʊ��������q�Ɠ]�ʗ}�����q��Lac �I�y�����߂��� p.435
- �ۈ�`�q�̒��ߒ���DNA �̃��[�v�`�����N���� p.437
- RNA �|�������[�[�̌݊����T�u���j�b�g��p���Ĉ�`�q�̓]�ʒ��߂������ p.438
- �^�j�����ł͈�`�q�̓]�ʂ߂��镡�G�ȃX�C�b�`���i�����Ă��� p.439
- �^�j�����̈�`�q���ߗ̈�́C�v�����[�^�[�ƒ���DNA �Ƃ���Ȃ� p.440
- �^�j�����̈�`�q�������^���p�N�́C�]�ʊJ�n���ʂɂ�����RNA p.441
- �|�������[�[�Ɠ]�ʊ�{���q�̏W���𑣐i���� p.441
- �^�j�����̈�`�q�������^���p�N�̓N���}�`���\�����Ǐ��I�ɕς��铭�������� p.442
- ��`�q�������^���p�N�͑���I�ɓ��� p.444
- �^�j�����̈�`�q�}���^���p�N�͂��܂��܂ȕ��@�œ]�ʂ�j�Q���� p.445
- �^�j�����̈�`�q���߃^���p�N�͂��������I��DNA �Ɍ�������@p.445
- �V���E�W���E�o�G�̔����ߒ��𐧌䂷�镡�G�Ȉ�`�q�X�C�b�`�͏����W���[������\�z����� p.447
- �V���E�W���E�o�G��Eve ��`�q�͑g�ݍ��킹�ɂ�钲�߂��Ă��� p.448
- �M���ނ̕��G�Ȉ�`�q���ߗ̈���P���Ȓ��߃��W���[������ł��Ă���@p.450
- �C���X���[�^�[�͐^�j�����̈�`�q���߃^���p�N�����ꂽ��`�q�ɂ���ڂ��e������������ p.452
- ��`�q�X�C�b�`�͋}���ɐi�����Ă��� p.453
- �܂Ƃ� p.454
���擪�s���E�I��\�ցA�͐߂̖ڎ��� �A�\�Q����
|
| 7��
�@�S�� |
7-4. ��剻�����זE�����o�����q��`�@�\�i17���ځj�@ p.454
- DNA �̍ĕҐ����W����ۂ̑��ψ� p.454
- ��A�̈�`�q���߃^���p�N�Q���o��y��̍זE�^�����肷�� p.455
- �������̍�����}������2 �̃^���p�N����λ�t�@�[�W�̍זE���ł̏�Ԃ����߂� p.457
- �P���Ȉ�`�q���߉�H��p���ċL�����u�����o���� p.458
- �]�ʉ�H�ɂ���čזE�͘_�����Z�𐋍s�ł��� p.459
- �\���I�����w�͊����̐��̕��i����V�������u��n�o���� p.460
- �T�����v�͈�`�q���߂̃t�B�[�h�o�b�N���[�v�Ɋ�Â��Ă��� p.460
- 1 �̈�`�q���߃^���p�N����A�̈�`�q�̔��������������邱�Ƃ��ł��� p.462
- ����I�Ȉ�`�q���߃^���p�N�̔������C�����ɂ����A�̈�`�q�S�̂̔�����U�����邱�Ƃ����� p.463
- �g�ݍ��킹�ɂ���`�q���߂��^�j�����̑��l�ȍזE�^������Ă��� p.464
- 1 �̈�`�q���߃^���p�N���튯�S�̂̌`���̈������ƂȂ肤�� p.465
- �Ғœ����̍זE������Ƃ�DNA �̃��`�����p�^�[�����p����� p.467
- �Q�m�����荞�݂�DNA �̃��`�����Ɋ�Â��Ă��� p.468
- �M���ނł́CCG �ɕx��“��”�ɑ����̈�`�q�����݂��� p.470
- �G�s�W�F�l�e�B�b�N�@�\�ɂ��C���肵����`�q�����̃p�^�[�������זE�ւƎp����� p.471
- ���F�̑S�̂ɍL�������N���}�`���\���̕ω����p����� p.473
- ��`�q�����̒��߂͖{���I�Ƀm�C�Y�ɗ������ p.476
- �܂Ƃ� p.477
���擪�s���E�I��\�ցA�͐߂̖ڎ��� �A�\�Q����
|
| 7��
�@�T�� |
7-5. �]�ʌ�̒����i19���ځj�@ p.477
- �]�ʂ̃A�e�j���G�[�V������RNA ���q�̍����𒆓r�ŏI��点�� p.477
- ���{�X�C�b�`�͑��Ì^�̈�`�q���߂�������Ȃ��@p.478
- �I��IRNA �X�v���C�V���O�ɂ��C�����`�q���畡���̕��q��̃^���p�N������邱�Ƃ��ł���@p.479
- �I��IRNA �X�v���C�V���O�̔����ɂ��C��`�q�̒�`�̏C�����K�v�ɂȂ����@p.480
- �V���E�W���E�o�G�̐�����́C��A��RNA �X�v���C�V���O�̒��߂ɂ��@p.481
- �]�ʎY��RNA �̐ؒf�E�|��A �t�����ʂ̕ω��ɂ��^���p�N����C ���[���ς��@p.482
- RNA �̕ҏW�ɂ��RNA �̂���`���̈Ӗ����ς��@p.483
- �j�����RNA �A�����������@p.485
- �זE���̓���̗̈�ɋǍ݂���mRNA p.486
- mRNA ��59 ����39 ���̔�|��̈悪�|��߂��Ă���@p.488
- �J�n���q�̃����_�����^���p�N������S�̂Ƃ��Đ��䂷��@p.488
- �^�j�����̖|��J�n�́C�|��J�n���ʂ̏㗬�ɂ���AUG �R�h���ł̊J�n�ɂ�蒲�߂ł���@p.489
- �z������̃��{�\�[���i�����ʂɂ��|��̒��߁@p.491
- mRNA ���萫�̕ω�����`�q�����𐧌䂷�邱�Ƃ�����@p.492
- �זE���ł̃|��A �̕t���ɂ��|��߂ł���@p.493
- �����q�̔�|��]�ʎY��RNA �����A���̑����̈�`�q�߂��Ă���@p.493
- RNA ���͍זE�̖h��@�\�ł���@p.495
- RNA ���ɂ��w�e���N���}�`�����`������邱�Ƃ�����@p.496
- RNA ���͋��͂Ȏ�������ɂȂ��Ă���@p.497
- �܂Ƃ߁@p.497
�͖����@p.497
���@���@p.499
���擪�s���E�I��\�ցA�͐߂̖ڎ��� �A�\�Q����
|
| ��V�� |
������@
�i2�� 7�� 76���� 114�Łj |
Part III |
Methods |
| �W�� |
�^���p�N���CDNA�CRNA�̑���
�i�\���F5�� 51���ځj |
Ch.8 |
Manipulating Proteins, DNA, and RNA |
| 8��
�@�P�� |
8-1. �זE�̒P���Ɣ|�{�i6���ځj�@ p.502
- �זE��g�D����P������@p.502
- �זE�͔|�{���邱�Ƃ��ł���@p.502
- ���������^�j�זE�͋ψ�ȍזE���Ƃ��čL���g���Ă���@p.505
- ���זE�͈�ÂɊv���������炷�@p.505
- �̍זE�j�̈ڐA�Ōl���Ƃɐ�p�̊��זE�����o�����Ƃ��ł���@p.507
- �n�C�u���h�[�}�זE�����g�����m�N���[�i���R�̂��ʂɐ��Y�ł���@p.508
- �܂Ƃ߁@p.510
���擪�s���E�I��\�ցA�͐߂̖ڎ��� �A�\�Q����
|
| 8��
�@�Q�� |
8-2. �^���p�N���̐����i6���ځj�@ p.510
- �זE���\�������ɕ��悷��@p.510_
- �זE�̋@�\��͂ɍזE���o�t���g���@p.511
- �^���p�N�����N���}�g�O���t�B�[�������@p.512
- �^���p�N���̓��ٓI�Ȍ������ʂ𗘗p����A�t�B�j�e�B�[�N���}�g�O���t�B�[�@p.513
- ��`�q����ɂ�镪�q�W���Ń^���p�N�������e�ՂɂȂ�@p.514
- �����������זE�n�ŕ��q�̋@�\���ɋᖡ����@p.516
- �܂Ƃ߁@p.516
���擪�s���E�I��\�ցA�͐߂̖ڎ��� �A�\�Q����
|
| 8��
�@�R�� |
8-3. �^���p�N���̉���i14���ځj�@ p.517
- �^���p�N��������SDS �|���A�N�����A�~�h�Q���d�C�j���@p.517
- ����̃^���p�N�����R�̂�p�����u���b�g�ɂ���Č��o����@p.518
- ���ʕ��͖@�ɂ�薢�m�^���p�N�̊��x�̍������肪�ł���@p.519
- ���̕����@�͓��Ɍ��ʓI�ł���@p.521
- ���̗͊w�����p���ă^���p�N�����̂̑傫����`��m��@p.522
- �����w�I��@�ő��ݍ�p���Ă���^���p�N���Q�肷��@p.523
- �^���p�N���ԑ��ݍ�p��2�n�C�u���b�h�@�ł�����ł���@p.523
- ���l�ȕ��@�œ���ꂽ�f�[�^��g�ݍ��킹�āC�^���p�N���ԑ��ݍ�p�̐M���x�̍����n�}�����@p.524
- ���w�I���@�ɂ��^���p�N���ԑ��ݍ�p�����A���^�C���Œ��ׂ�@p.524
- �P��̕��q���ώ@����Z�p������@p.526
- �^���p�N���̋@�\�������q�ɂ���đI��I�ɑj�Q����@p.527
- �^���p�N���̍\�������X ����܂��g���@p.527
- NMR �ŗn�t���̃^���p�N���̍\��������ł���@p.529
- �^���p�N���̃A�~�m�_�z��ƍ\������@�\�̃q���g��������@p.530
- �܂Ƃ߁@p.532
���擪�s���E�I��\�ցA�͐߂̖ڎ��� �A�\�Q����
|
| 8��
�@�S�� |
8-4. DNA�̉�͂Ƒ����i15���ځj�@ p.532
- �傫��DNA ���q�𐧌��y�f�Œf�Ђɂ���@p.532
- �Q���d�C�j���ő傫���̈قȂ�DNA ���q������@p.534
- ���ː����ʑ̂�W���������g���Đ���DNA ���q��in vitro �ŕW���ł���@p.535
- �n�C�u���b�h�`���ɂ��j�_�̓���̉���z������x�悭���o����@p.535
- �m�U���E�u���b�g�C�T�U���E�u���b�g�ɂ��C�d�C�j���ŕ��������j�_���q�̃n�C�u���b�h�`���ׂ�@p.538
- DNA ���C�u�����[������`�q���N���[�j���O����@p.540
- �ʁX�̖ړI�Ɏg��2 ��ނ�DNA ���C�u�����[�@p.541
- cDNA �N���[���̖|��̈�͕��f����Ă��Ȃ��@p.544
- PCR �����ɂ��D�݂̈�`�q���ł���@p.544
- �זE��]�݂̃^���p�N���̐����H��Ƃ��ė��p����@p.546
- �^���p�N����j�_�����w�����ɂ���Ē��ڍ�������@p.548
- DNA �̉���z��͂��₭����ł���@p.548
- ����z��^���p�N���̃A�~�m�_�z��𐄒肷��@p.550
- ���l�Ȑ����̑S�Q�m������z���肳��Ă���@p.551
- �܂Ƃ߁@p.552
���擪�s���E�I��\�ցA�͐߂̖ڎ��� �A�\�Q����
|
| 8��
�@�T�� |
8-5. ��`�q�̔����Ƌ@�\�̉���i20���ځj�@ p.553
- �ÓT��`�w�ł̓����_���ɕψق�U�����זE�̋@�\�Ȃ������̂�Ƃ��납�猤�����n�߂�@p.553
- ��`�q�X�N���[�j���O�ɂ��ُ�̂���ψّ̂肷��@p.556
- �ψقɂ��^���p�N���̋@�\�͌������C���邢�͐V�@�\���l������@p.557
- ��������ɂ��C2 �̕ψق�������`�q�ɐ������̂��Ⴄ��`�q�ɐ������̂��ʂ���@p.558
- �����o�H�œ�����`�q�̏�������ʉ�͂ɂ�茈�߂邱�Ƃ��ł���@p.558
- �ψقɂ�蓯�肳�ꂽ��`�q�̈ʒu�����߂�@p.559
- �q�g�̈�`�w�����͓��L�̖����͂��ł��邪�傫�ȉ\�������@p.560
- �q�g�̈�`�q�̓n�v���^�C�v�u���b�N�ň�`���C����͕a���ƂȂ�ψق̒T���ɖ𗧂@p.561
- ���G�Ȍ`���͕����̈�`�q�̉e������@p.563
- �t��`�w�͊��m�̈�`�q����n�߂āC���̋@�\���זE�̂ǂ̔����ɕK�v�������肷��@p.563
- ��`�q�͂��낢��Ȃ����ʼn��ςł���@p.564
- ���ψ�`�q�͂��낢��Ȑ����̍זE�ɑ}���ł���@p.565
- ��������`�q����ɂ����ς���@p.566
- ��`�q�����A���́C�זE�����w�ɂƂ��Ă��_�ƂɂƂ��Ă��d�v�ł���@p.568
- �W�����m�b�N�A�E�g�ψّ̂��K�͂Ɏ��W���đS��`�q�̋@�\�ׂ��i�Ƃ���@p.569
- RNA ���ɂ���Ĉ�`�q�@�\���ȒP�v���ɂ킩��@p.571
- ���|�[�^�[��`�q��in situ �n�C�u���b�h�`���@�ň�`�q�̔������ʁC���������𖾂炩�ɂ���@p.572
- �X�̈�`�q�̔������ʓI�t�]��PCR ��p���đ��肷��@p.573
- �}�C�N���A���C�Ő�����̈�`�q�̔�������x�ɒǐՂł���@p.574
- �P��זE�̈�`�q��������͂���Ɛ����w�I“�m�C�Y”���킩��@p.575
- �܂Ƃ߁@p.576
�͖����@p.576
���@���@p.578
���擪�s���E�I��\�ցA�͐߂̖ڎ��� �A�\�Q����
|
| �X�� |
�זE�̊ώ@
�i�\���F2�� 25���ځj |
Ch.9 |
Visualizing Cells |
| 9��
�@�P�� |
9-1. ���w�������ōזE�������i17���ځj�@ p.579
- ���w�������̕���\�̌��E��0.2 mm �ł���@p.580
- �ʑ�������������������������g���Ɛ������זE���͂����茩����@p.583
- �f�W�^���Z�p��p���ĉ摜�̑����Ɖ�͂��ł���@p.583
- �������ώ@�ɂ͑g�D���Œ肵�ؕЂ��쐻����@p.585
- ����̕��q�̍זE�����z�͌u���������ł킩��@p.586
- ����̕��q�̌��o�ɍR�̂����p�ł���@p.588
- ���w�������ł����G�ȎO�����\���̉摜��������悤�ɂȂ����@p.589
- ���œ_�����^�������͏œ_�̂��ꂽ����r���������w�ɂ��ؕЂ����@p.590
- �u���^���p�N�𗘗p���āC�זE����̂̃^���p�N�������܂܌ʂɕW���ł���@p.592
- ���זE���̃^���p�N���̓������ǐՂł���@p.593
- �}���ɕω�����זE���C�I���Z�x���w����ő��肷��@p.596
- ���߂��Ȃ��������זE�֓���������@������������@p.597
- ���͉摜�邾���łȂ��C�������̂̑���ɂ��𗧂@p.598
- �S���ˏƖ��u���������@�ŌX�̕��q��������@p.599
- ���q�ԗ͌������@�ŌX�̕��q��G���ē�������@p.600
- ���q����ː����ʑ̂ŕW������@p.600
- ���ː����ʑ̂�p���čזE����̓��̕��q��ǐՂ���@p.602
- �܂Ƃ߁@p.603
���擪�s���E�I��\�ցA�͐߂̖ڎ��� �A�\�Q����
|
| 9��
�@�Q�� |
9-2. �d�q�������ōזE�ƕ��q�������i8���ځj�@ p.604
- �d�q�������͍זE���̔��\���������������@p.604
- �d�q�������p�̐��������ɂ͓��ʂ̒����@���K�v�ł���@p.605
- �Ɖu���d�q�������œ���̋��啪�q�̈ʒu���킩��@p.606
- �����^�d�q�������ł͕\�ʂ̎O��������������@p.607
- �����V���h�E�C���O��p����ƁC���ߌ^�d�q�������ŕ\�ʂ̂悤�������{���Œ��ׂ���@p.608
- �l�K�e�B�u���F�@�ł��ቷ�d�q�������@�ł�������\�ŋ��啪�q�̑���������@p.610
- �����̉摜�̑g�ݍ��킹�ŕ���\��ł���@p.610
- ���܂��܂ȕ�������Ȃ��߂��}��g�ݍ��킹�ĎO���������\�z����@p.612
- �܂Ƃ߁@p.612
�͖����@p.614
���@���@p.615
���擪�s���E�I��\�ցA�͐߂̖ڎ��� �A�\�Q����
|
| ��W�� |
�זE�̓����\��
�i9�� 38�� 389���� 483�Łj
|
Part IV |
Internal Organization of the Cell |
| 10�� |
���̍\��
�i�\���F2��20���ځj |
Ch.10 |
Membrane Structure |
| 10��
�@�P�� |
10-1. ������d�w�i8���ځj�@ p.617
- �O���Z����������, �X�t�B���S����, �X�e���[���͍זE���̂����Ȏ����ł���@p.618
- ���������͎��R�ɓ�d�w���`������@p.620
- ������d�w�͓��̗����̂ł���@p.621
- ������d�w�̗������́C���̍\�������ɂ���Č��܂�@p.622
- ������d�w�͗�����������ɂ�������炸�C�g���̈قȂ�������@p.624
- ���b�H�̓��������P���q�w�Ɉ͂܂�Ă���@p.625
- ������d�w�̔�Ώ̍\���͋@�\��d�v�ł���@p.626
- ���ׂĂ̍זE���̕\�ʂɂ͓����������݂���@p.628
- �܂Ƃ� p.629
���擪�s���E�I��\�ցA�͐߂̖ڎ��� �A�\�Q����
|
| 10��
�@�Q�� |
10-2. ���^���p�N�i12���ځj�@ p.629
- ���^���p�N�͂��܂��܂ȕ��@�Ŏ�����d�w�ɑg�ݍ��܂�Ă���@p.629
- �����A���J�[�̓V�O�i���^���p�N�̖��ւ̌����𐧌䂷��@p.630
- �����̖��ђʃ^���p�N�ł̓|���y�v�`�h����α�w���b�N�X������������d�w���т��Ă���@p.631
- ���ђ�α�w���b�N�X�͂������ɑ��ݍ�p���邱�Ƃ������@p.632
- β�o�����̂Ȃ��ɂ͖����ђʂ����^�̃`���l���������̂�����@p.634
- ���^���p�N�̑����ɂ͓��������Ă���@p.635
- ���^���p�N�́C�E�ʊ����܂�p���ĉn���E�����ł���@p.636
- �o�N�e���I���h�v�V���͎�����d�w���ђʂ���7 �{��α�w���b�N�X����Ȃ�C���쓮�v���g���|���v�Ƃ��ē����@p.640
- ���^���p�N�͑�^�̕����̂Ƃ��ċ@�\���Ă��邱�Ƃ������@p.642
- ���^���p�N�̑����͖����ʓ����g�U����@p.642
- �זE�̓^���p�N���⎉�����C���̓�������ɋǍ݂����邱�Ƃ��ł���@p.645
- ��w�̍זE���i�͍זE���ɋ@�B�I���x��^���C���^���p�N�̊g�U����������@p.646
- �܂Ƃ߁@p.648
�͖����@p.648
���@���@p.650
���擪�s���E�I��\�ցA�͐߂̖ڎ��� �A�\�Q����
|
| 11�� |
�����q�̖��A���Ɩ��̓d�C�I����
�i�\���F4�� 27���ځj |
Ch.11 |
Membrane Transport of Small Molecules and the Electrical Properties of Membranes |
| 11��
�@�P�� |
11-1. ���A���̊�{�i3���ځj�@ p.651
- �^���p�N�����܂܂Ȃ�������d�w�̓C�I�����قƂ�ǒʂ��Ȃ��@p.652
- ���ɂ����v�ȗA���^���p�N�́C�A���̂ƃ`���l����2��ނł���@p.652
- �A���̂��s���\���A���̓G�l���M�[���Ƌ������Ă���@p.653
- �܂Ƃ߁@p.654
���擪�s���E�I��\�ցA�͐߂̖ڎ��� �A�\�Q����
|
| 11��
�@�Q�� |
11-2. �A���̂Ɣ\�����A���i7���ځj�@ p.654
- �C�I�����z�ɂ��\���A���@p.656
- �זE���ɂ���A���̂��זE���]����pH �߂��Ă���@p.657
- ���זE�ł͗A���̂���Ώ̓I�ɕ��z���āC�n�����זE���f�A������@p.658
- ATP �쓮�^�|���v��3 ��ނ���@p.659
- Ca2+�|���v��P �^ATP �A�[�[�̂����ōł��������i��ł���@p.660
- �זE����P �^Na�{–K�{�|���v�͖����͂���Na+���z�����o���@p.661
- ABC �A���͖̂��A���^���p�N�̍ő�̃t�@�~���[���Ȃ��@p.663
- �܂Ƃ߁@p.667
���擪�s���E�I��\�ցA�͐߂̖ڎ��� �A�\�Q����
|
| 11��
�@�R�� |
11-3. �C�I���`���l���Ɩ��̓d�C�I�����i3���ځj�@ p.667
- �C�I���`���l���ɂ͑I�𐫂�����C�J��Ԃƕ�Ԃ����݂ɌJ��Ԃ��@p.667
- �����זE�����d�ʂ͂����ɁCK�{�R�k�`���l���ƍזE�����͂���K�{�̌��z�ɂ���Đ�����@p.669
- �Î~���d�ʂ́CNa�{–K�{�|���v����~����Ƃ������Ə��ł���@p.669
���擪�s���E�I��\�ցA�͐߂̖ڎ��� �A�\�Q����
|
| 11��
�@�S�� |
11-4. �ۂ�K1 �`���l���̎O�����\������C�C�I���`���l���̓��������킩���i14���ځj�@ p.671
- �A�N�A�|�����͐���ʂ����C�I���͒ʂ��Ȃ��@p.673
- �j���[�����̋@�\�͒����̂т��\���Ɉˑ����Ă���@p.675
- �d�C�I�������������זE�ł͓d�ʈˑ��z�C�I���`���l���������d�ʂ�������@p.676
- �~�G�������́C�_�o�זE�ł̊����d�ʂ̓`�B���x�ƌ��������߂Ă���@p.678
- �p�b�`�N�����v�@�ɂ��C�X�̓d�ʈˑ��`���l���̓I���E�I�t�^�̕����ŊJ���Ă��邱�Ƃ��킩��@p.680
- �d�ʈˑ��z�C�I���`���l���́C�i���I�ɂ��\���I�ɂ����݂Ɋ֘A���Ă���@p.682
- ���w�V�i�v�X�œ`�B�����ˑ��C�I���`���l�������w�V�O�i����d�C�V�O�i���ɕϊ�����@p.682
- ���w�V�i�v�X�͋������ɂ��}�����ɂ��Ȃ�@p.684
- �_�o�ؐڍ����̃A�Z�`���R������e�͓̂`�B�����ˑ��z�C�I���`���l���ł���@p.684
- �`�B�����ˑ��C�I���`���l���͌����_��̎�v�W�I�ł���@p.686
- �_�o�Ƌ̓`�B�ł�5 ��ނ̃C�I���`���l������������������@p.687
- �j���[�����͕��G�ȉ��Z�������u�ł���@p.688
- �j���[�����ł̏���ɂ́C���Ȃ��Ƃ�3 ��ނ�K1 �`���l���̘A�g���K�v�ł���@p.689
- �M���ނ̊C�n�ł̒��������iLTP�j�́CNMDA ��e�̃`���l����ʂ�Ca2+ �̗����Ɉˑ����Ă���@p.691
- �܂Ƃ߁@p.692
�͖����@p.693
���@���@p.694
���擪�s���E�I��\�ցA�͐߂̖ڎ��� �A�\�Q����
|
| 12�� |
�זE�����ƃ^���p�N���̑I��
�i�\���F5�� 34���ځj �@
|
Ch.12 |
Intracellular Compartments and Protein Sorting |
| 12��
�@�P�� |
12-1. �זE�̋�扻�i5���ځj�@�@ p.695
- �^�j�זE�ɂ͕K���C���܂����g�ݍ��킹�̖��ň͂܂ꂽ���튯������@p.695
- �e���튯�̈ʒu�W�́C�i����̋N�����l����Ɛ����ł���@p.697
- �^���p�N���������̊Ԃ��ړ�������@�͂��܂��܂ł���@p.699
- �V�O�i���z��̓^���p�N�����זE�̓K�ȏꏊ�ɗU������@p.701
- ���튯�̑唼�̓[������̍������s�\�ŁC���튯���g�̂�����K�v�Ƃ���@p.702
- �܂Ƃ߁@p.704
���擪�s���E�I��\�ցA�͐߂̖ڎ��� �A�\�Q����
|
| 12��
�@�Q�� |
12-2. �j�|�זE���Ԃ̕��q�̗A���i7���ځj�@ p.704
- �j���E�����̂͊j�����ђʂ��Ă���@p.705
- �j�^���p�N�͊j�Ǎ݉��V�O�i���ɂ���Ċj�ɗU�������@p.705
- �j��������e�̂͊j�Ǎ݉��V�O�i����NPC �^���p�N�̗����Ɍ�������@p.707
- �j�O�ւ̔��o�͊j���ւ̎�荞�݂ƕ������t�Ȃ����Ŏ��Ă���@p.708
- Rao GTP �A�[�[��NPC ��ʂ�A���̕����������߂�@p.708
- NPC ��ʂ�A���̒��߂́C�A�����u�֕������߂Â��Ȃ��Ƃ������@�ɂ��@p.709
- �j���͗L������̍ۂɂ��ɂȂ�@p.710
- �܂Ƃ߁@p.712
���擪�s���E�I��\�ցA�͐߂̖ڎ��� �A�\�Q����
|
| 12��
�@�R�� |
12-3. �~�g�R���h���A�Ɨt�Α̂ւ̃^���p�N�A���i6���ځj�@ p.713
- �~�g�R���h���A�ւ̓]���́C�V�O�i���z��ƃ^���p�N�]�����u�Ɉˑ�����@p.713
- �~�g�R���h���A�̃^���p�N�O��̂́C�قǂ����|���y�v�`�h���̏�ԂŎ�荞�܂��@p.715
- �}�g���b�N�X�ւ̃^���p�N���̎�荞�݂́CATP �̉��������Ɩ��d�ʂɂ���ċ쓮�����@p.716
- �ۂƃ~�g�R���h���A�ł̓|�������O���ɑ}�����邵���݂����Ă���@p.717
- �~�g�R���h���A�����Ɩ��ԍo�ւ̗A���ɂ͐���ނ̌o�H������@p.717
- �t�Α̂̃`���R�C�h���Ƀ^���p�N����A������ɂ�2 ��ނ̃V�O�i���z�K�v�ł���@p.719
- �܂Ƃ߁@p.720
���擪�s���E�I��\�ցA�͐߂̖ڎ��� �A�\�Q����
|
| 12��
�@�S�� |
12-4. �y���I�L�V�\�[���i2���ځj 721
- �y���I�L�V�\�[���͕��q��_�f�Ɖߎ_�����f���g���Ď_���������s���@p.721
- �^���p�N���̃y���I�L�V�\�[���ւ̎�荞�݂͒Z���V�O�i���z�w������@p.722
- �܂Ƃ߁@p.723
���擪�s���E�I��\�ցA�͐߂̖ڎ��� �A�\�Q����
|
| 12��
�@�T�� |
12-5. ���E���i14���ځj�@ p.723
- ���E�͍̂\�����@�\�����܂��܂ł��� p.724
- �V�O�i���z��͑e�ʏ��E�̂Ɏ�荞�܂��^���p�N���ōŏ��Ɍ������� p.726
- �V�O�i�����ʗ��q�iSRP�j�����E�̃V�O�i���z���e�ʏ��E�̖��ɂ�����ٓI��e�̂֓����@p.727
- �|���y�v�`�h���͓]�����u�̐��̒ʂ鏬�E��ʉ߂��� p.730
- ���E�̖���ʉ߂���]���ɂ́C�|���y�v�`�h���̓��������͕s���ł͂Ȃ� p.731
- �P�ђʃ^���p�N�ł́C�|���y�v�`�h���̓r���ɂ��鏬�E�̃V�O�i���z�����ђʂ���α�w���b�N�X�̌`�Ŏ�����d�w���ɂƂǂ܂� p.732
- �����ђʃ^���p�N�̗��̍\���͗A���J�n�ƒ�~���w������V�O�i���̑g�ݍ��킹�ɂ�茈�܂� p.734
- �]�����I������|���y�v�`�h���͑e�ʏ��E�̓��o�Ő܂肽���܂�g�ݗ��Ă���@p.736
- �e�ʏ��E�̂ō����^���p�N���̑唼�ɂ͋��ʂ�N�|�����^�I���S�����t�������@p.736
- �I���S���̓^���p�N���̐܂肽���ݏ�Ԃ������W���Ƃ��Ďg����@p.738
- �܂肽���݂�������^���p�N���͏��E�̂���^�яo����čזE���]���ŕ��������@p.739
- ���E�̂ɂ���܂肽���݂�������^���p�N���͕ϐ��^���p�N����������������@p.740
- ���^���p�N�̂Ȃ��ɂ́C�O���R�V���z�X�t�@�`�W���C�m�V�g�[���iGPI�j�A���J�[�����L�������Ă�����̂�����@p.742
- ���̎�����d�w�̑唼�͏��E�̂őg�ݗ��Ă���@p.743
- �܂Ƃ߁@p.745
�͖����@p.746
���@���@p.748
���擪�s���E�I��\�ցA�͐߂̖ڎ��� �A�\�Q����
|
| 13�� |
�זE���ɂ����鏬�E�̈ړ�
�i�\���F5�� 51���ځj |
Ch.13 |
Intracellular Vesicular Traffic |
| 13��
�@�P�� |
13-1. ���A���̕��q�@�\�ƍזE�����̑��l���ێ��i11���ځj�@ p.750
- �핢���E�ɂ͂��낢��Ȏ�ނ�����@p.751
- �N���X�����핢�̌`���ŏ��E���ł���@p.754
- ������\�������Ȃ��핢������@p.755
- �z�X�z�C�m�V�`�h�͍זE���튯�▌���̖ڈ�ƂȂ�@p.757
- �זE���^���p�N���핢���E�̖�����̐藣���ƒE�핢�߂���@p.757
- �P�ʑ�GTP �A�[�[���핢�̑g�ݗ��Ă߂���@p.758
- �A�����E�͂��ׂĂ����`�Ƃ����킯�ł͂Ȃ��@p.760
- Rab �^���p�N�͏��E��W�I�ɓ����@p.760
- SNARE �����̗Z���𒇉��@p.762
- ������Ԃɂ���SNARE ���ӂ����ѓ�������ɂ́C�𗣂��K�v�ł���@p.764
- �E�C���X�̗Z���^���p�N��SNARE �͓��l�ȗZ���@�\���g���Ă���@p.764
- �܂Ƃ߁@p.766
���擪�s���E�I��\�ցA�͐߂̖ڎ��� �A�\�Q����
|
| 13��
�@�Q�� |
13-2. ���E�̂���S���W�̂��o�R����A���i11���ځj�@�@ p.766
- �^���p�N����COPII �핢�A�����E�ɓ����ď��E�̂��瑗��o�����@p.767
- �������܂肽���݂ƏW����Ԃ̃^���p�N�����������E�̂�����o�����@p.767
- ���E�̂���S���W�̂ւ̗A���͏��E���ǃN���X�^�[�ɂ���čs����@p.768
- ���E�̂ւ̉���ߒ��͑I�ʃV�O�i����p���čs����@p.769
- �^���p�N���̑����́C�����ׂ����ɑI��I�ɕۗ������@p.771
- �S���W�̂͐��R�Ƃ�����A�̋�悩��Ȃ�@p.771
- �I���S�����̓S���W�̂ʼn��H�����@p.773_
- �v���e�I�O���J���̓S���W�̂ō��グ����@p.775
- �����t���͉��̂��߂ɍs����̂��낤�@p.776
- �S���W�̂�ʉ߂���A���́C���E�A�����邢�͔X���n�ɂ���čs����@p.777
- �}�g���b�N�X�^���p�N�̓S���W�w�̑g�D����������@p.778
- �܂Ƃ߁@p.779
���擪�s���E�I��\�ցA�͐߂̖ڎ��� �A�\�Q�������擪�s���E�I��\�ցA�͐߂̖ڎ��� �A�\�Q����
|
| 13��
�@�R�� |
13-3. �g�����X�S���W�Ԃ��烊�\�\�[���ւ̗A���i10���ځj�@ p.779
- ���\�\�[���͍זE���ŏ������s����v���ʂł���@p.779
- ���\�\�[���͕s�ψ�ȏ��튯�ł���@p.780
- �A����ۗނ̉t�E�͍ۗ����đ��l�Ȗ����������\�\�[���ł���@p.781
- ���\�\�[���ɕ����𑗂荞�ތo�H�͂���������@p.782
- �}���m�[�X6�|�����_��e�̂̓g�����X�S���W�ԂŃ��\�\�[���^���p�N�����ʂ���@p.783
- �}���m�[�X6�|�����_��e�͓̂���̖��̊Ԃ���������@p.784
- ���������y�f�̃|���y�v�`�h���ɂ���V�O�i���p�b�`��M6P �̕t���̎肪����ƂȂ�@p.785
- �q�g��GlcNAc �����_��]�ڍy�f�Ɍ��ׂ�����ƁC���\�\�[���~�Ϗǂ������N�������@p.785
- ���\�\�[���̂Ȃ��ɂ̓G�L�\�T�C�g�[�V�X�������̂�����@p.786
- �܂Ƃ߁@p.787
���擪�s���E�I��\�ցA�͐߂̖ڎ��� �A�\�Q�������擪�s���E�I��\�ցA�͐߂̖ڎ��� �A�\�Q����
|
| 13��
�@�S�� |
13-4. �זE������זE���ւ̗A�� �� �G���h�T�C�g�[�V�X�i9���ځj�@ p.787
- ���̐H�זE�͑傫�ȗ��q��ێ�ł���@p.787
- ����p���E�͍זE���̔핢�s�b�g������������@p.789
- ����p���E�̓N���X�����핢�����ł͂Ȃ��@p.790
- �זE�͎�e�̂�����G���h�T�C�g�[�V�X�ɂ��O���狐�啪�q����荞�ށ@p.791
- �G���h�T�C�g�[�V�X�Ŏ�荞�܂ꂽ�����̂����G���h�\�[������������Ȃ����̂́C���\�\�[���ɂ܂킳���@p.792
- ����̃^���p�N���������G���h�\�[������������čזE���ɕԑ������@p.793
- ����G���h�\�[���������o�H�ɑ��E�̂��`�������@p.795
- ���啪�q�͏��זE�w���g�����X�T�C�g�[�V�X�ɂ���Ēʉ߂���@p.797
- ���זE�ɂ�2 ��ނ̏����G���h�\�[����悪���邪�C����G���h�\�[�����͋��ʂ��Ă���@p.798
- �܂Ƃ߁@p.798
���擪�s���E�I��\�ցA�͐߂̖ڎ��� �A�\�Q����
|
| 13��
�@�T�� |
13-5. �g�����X�S���W�Ԃ���זE�O���ւ̗A�� �� �G�L�\�T�C�g�[�V�X�i10���ځj 799
- �^���p�N���⎉���̑����̓S���W�̂���זE�\�ʂ֎����I�ɗA�������炵���@p.800
- ���召�E�̓g�����X�S���W�Ԃ���o�肷��@p.801
- �^���p�N���͕��召�E�̌`���ߒ��Ń^���p�N�����̉ߒ����o�ĉ��H�����@p.803
- ���召�E�͓��e����o������V�O�i��������܂ōזE���̋߂��őҋ@���Ă���@p.803
- ���ߐ��G�L�\�T�C�g�[�V�X�́C����ꂽ�͈͂̍זE���Ƃ��̒����̍זE�����֗^����Ǐ��I�ȉ����ł���@p.804
- ���召�E�̖������͍זE�����炷�₭��菜�����@p.805
- ���ߐ��G�L�\�T�C�g�[�V�X�ɂ́C�זE�����L�������������@p.805
- �ɐ��̂���זE�ł́C�g�����X�S���W�ԗR���̃^���p�N���͍זE���̌��܂������ɗA�������@p.805
- ���^���p�N�Ǝ�����I��I�ɐ������זE�����֓������낢��ȕ����@p.806
- �V�i�v�X���E�̓G���h�T�C�g�[�V�X���E���璼�ڌ`�������@p.807
- �܂Ƃ߁@p.809
�͖����@p.810
���@���@p.812
���擪�s���E�I��\�ցA�͐߂̖ڎ��� �A�\�Q����
|
| 14�� |
�G�l���M�[�ϊ�
�F�~�g�R���h���A�Ɨt�Α��i�\���F5�� 71���ځj |
Ch.14 |
Energy Conversion
: Mitochondria and Chloroplasts |
| 14��
�@�P�� |
14-1. �~�g�R���h���A�i11���ځj�@ p.815
- �~�g�R���h���A�ɂ͊O���Ɠ���������C������2 �̋��ɕ�����Ă���@p.816
- �N�G���_��H�͍��G�l���M�[�d�q�ݏo���@p.817
- ���w�Z����p���_�������̃G�l���M�[��ATP �ɕϊ�����@p.817
- �d�q��3 �̑傫�Ȍċz�y�f�����̂��o��NADH ����_�f�ɓ`�B�����@p.819
- �d�q���ċz�����ړ�����ۂɕ��o�����G�l���M�[�́C�~�g�R���h���A�������͂��ޓd�C���w�I�ȃv���g�����z�̂������Œ~������@p.820
- �v���g�����z��ATP ������i�߂�@p.821
- �v���g�����z�����������鋤��A�����쓮����@p.822
- �זE��ATP �̂قƂ�ǂ̓v���g�����z���琶�Y�����@p.822
- �~�g�R���h���A�͍זE����ATP/ADP �̔�������l�ɕۂ��Ă���@p.823
- �זE��ATP �𗘗p����ɂ́CATP ����������DG ���傫�ȕ��̒l�ɂȂ��Ă��Ȃ���Ȃ�Ȃ��@p.824
- ATP �����y�f�͋t�����ɓ����āCATP ��������������H1 �����ݏo�����Ƃ�����@p.826
- �܂Ƃ߁@p.827
���擪�s���E�I��\�ցA�͐߂̖ڎ��� �A�\�Q�������擪�s���E�I��\�ցA�͐߂̖ڎ��� �A�\�Q����
|
| 14��
�@�Q�� |
14-2. �d�q�`�B�n�ƃv���g���|���v�i14���ځj�@ p.827
- �v���g���͂���߂Ĉړ����₷���@p.827
- �_���Ҍ��d�ʂ͓d�q�ɑ���e�a���������Ă���@p.828
- �d�q�`�B�ɂ���ʂ̃G�l���M�[�����o�����@p.829
- �ċz���̑����̓d�q�`�B�͕̂����w�I���@�œ��肳�ꂽ�@p.829
- �ċz���ɂ͓����ɖ��܂���3 �̑傫�ȍy�f�����̂����݂���@p.831
- �V�g�N�����_���y�f�̓S�|�����S��O2 �̊Ҍ��������I�ɐG�}����@p.832
- �d�q�`�B�́C�~�g�R���h���A�������ł̃����_���ȏՓ˂̍ۂ̓d�q�̃g���l�����ʂɂ���ċN����@p.834
- H�{�̂��ݏo���ɕK�v�ȃG�l���M�[�́C3 ��ނ̌ċz�y�f�����̊Ԃő傫���ω�����_���Ҍ��d�ʂ��瓾����@p.835
- H�{�|���v�̍�p�@�\�͎�v�ȍy�f������3 ��ނňقȂ�@p.835
- H�{�C�I�m�t�H�A�͓d�q�`�B��ATP ������E����������@p.836
- �ċz���߂͐��펞�Ɍċz���𗬂��d�q�̗ʂ�}������@p.837
- �V�R�̒E�����܂����F���b�זE���̃~�g�R���h���A�M���u�ɕς��Ă���@p.838
- �~�g�R���h���A�͍זE�̑�ӂ̈ێ��ɑ����̏d�v�Ȗ��������@p.838
- �ۂ��G�l���M�[�̗��p�ɉ��w�Z���@�\��p���Ă���@p.839
- �܂Ƃ߁@p.840
���擪�s���E�I��\�ցA�͐߂̖ڎ��� �A�\�Q����
|
| 14��
�@�R�� |
14-3. �t�Α̂ƌ������i16���ځj�@ p.840
- �t�Α͍̂זE���튯�ł���F�f�̂ɑ����Ă���@p.841
- �t�Α̂̓~�g�R���h���A�Ɏ��Ă��邪�C��悪1 �����@p.842_
- �t�Α̂͑��z����ߑ����C������g���ĒY�f�Œ������@p.843
- �Y�f�Œ�̓��u���[�X�r�X�����_�J���{�L�V���[�[�ɂ���ĐG�}�����@p.844
- 1 ���q��CO2 �Œ�ɂ�����ATP 3 ���q��NADPH 2 ���q��������@p.845
- �����̐A���ł͒�Z�x��CO2 �ł������ł���悤�ɒY�f�Œ蔽������扻����Ă���@p.846
- �������̓N�����t�B�����q�̍s�������w�����Ɉˑ�����@p.847
- �����w�n�͔������S�ƃA���e�i�����̂���\�z����Ă���@p.848
- �������S�ł́C�N�����t�B�����ߑ��������̃G�l���M�[�ɂ���Ďア�d�q���^�̂��狭�͂ȓd�q���^�̂������@p.849
- ��z�I�������_�������ł�NADPH ��ATP �����Y�����@p.850
- �t�Α̂͏z�I�������_�������ɂ���āCNADPH �̐����Ȃ���ATP �Y�ł���@p.853
- �����w�nI ��II �̍\���ɂ͊֘A��������C�ۂ̌����w�n�Ƃ����Ă���@p.853
- �v���g���쓮�͂̓~�g�R���h���A�Ɨt�Α̂œ����ł���@p.853
- �t�Α̂̓����ɂ���A���̃^���p�N�́C�זE���]���Ƃ̑�ӎY���̌����������ǂ�@p.854
- �t�Α̂͂ق��̏d�v�Ȑ��������s���Ă���@p.855
- �܂Ƃ߁@p.855
���擪�s���E�I��\�ցA�͐߂̖ڎ��� �A�\�Q����
|
| 14��
�@�S�� |
14-4. �~�g�R���h���A�ƐF�f�̂̈�`�q�n�i16���ځj�@ p.855
- �~�g�R���h���A�Ɨt�Α̂ɂ͊��S�Ȉ�`�q�n������@p.856
- �זE���̃~�g�R���h���A��F�f�̂̐��́C���튯�̐����ƕ���ɂ����ɕۂ����@p.857
- �~�g�R���h���A�Ɨt�Α̂̃Q�m���͑��l�ł���@p.859
- �~�g�R���h���A�Ɨt�Α̂͂Ƃ��ɍזE�ɋ��������ۂ��i���������̂炵���@p.859
- �~�g�R���h���A�̃R�h���g�p�͎��R�x�������C�قȂ��`�Í�������@p.861
- �����̃~�g�R���h���A�̈�`�q�n�́C���m�̂��̂̂Ȃ��ōł��ȒP�ł���@p.862
- ���튯�̈�`�q�ɂ̓C���g�������܂ނ��̂�����@p.863
- �����A���̗t�Α̃Q�m���ɂ͖�120 �̈�`�q������@p.863
- �~�g�R���h���A��`�q�͔��f����`�ɂ��p�����@p.864
- �����̐����ōזE���튯�̈�`�q�͕ꐫ��`�Ŏp�����@p.866
- �y��̃v�`�ψي��́C�~�g�R���h���A�̌`���ɍזE�j�����ɏd�v�Ȃ��Ƃ������Ă���@p.866
- �~�g�R���h���A�Ɨt�Α͍̂זE�j�Ɏw�߂���Ă���g�D���ٓI�^���p�N���܂ށ@p.867
- �~�g�R���h���A�͎��g�ɕK�v�Ȏ����̑啔�����O�������荞�ނ��C�t�Α̂͂��̑唼���������Ă���@p.867
- �~�g�R���h���A�͍זE��̘̂V���ɂ�������Ă���炵���@p.868
- �~�g�R���h���A�Ɨt�Α̂ɂ͂Ȃ��Ǝ��̈�`�q�n������̂��@p.868
- �܂Ƃ߁@p.870
���擪�s���E�I��\�ցA�͐߂̖ڎ��� �A�\�Q����
|
| 14��
�@�T�� |
14-5. �d�q�`�B�n�̐i���i15���ځj�@ p.870
- ���������̍זE�͂����炭���y�ɂ����ATP �Y���Ă����@p.870
- ���C���ۂ͓d�q�`�B�n�ɂ���āC�y���̕��q�������ȃG�l���M�[���Ƃ��ė��p�ł���悤�ɂȂ����@p.871
- ���s���̊Ҍ��͂āC�������ۂ͍זE�i���ɂ�����d��ȏ�Q�����������@p.872
- �V�A�m�o�N�e���A�̌������d�q�`�B�n�ɂ���đ�C���Ɏ_�f�������C�V���������`�Ԃ����܂ꂽ�@p.873
- �܂Ƃ߁@p.875
�͖����@p.877
���@���@p.878
���擪�s���E�I��\�ցA�͐߂̖ڎ��� �A�\�Q����
|
| 15�� |
�זE�̏��`�B�@
�i�\���F5�� 60���ځj
|
Ch.15 |
Mechanisms of Cell Communication |
| 15��
�@�P�� |
15-1. �זE�ɂ�������`�B�̊�{�i18���ځj�@ p.879
�@�i�����̉����͂��̃e�L�X�g�̂��ߕt�L�������́B�����ɂ͂Ȃ���j�@
- �זE�O�V�O�i�����q�͓��ٓI�Ȏ�e�̂Ɍ�������@p.880
- �זE�O�V�O�i�����q�́C�Z�����ł��������ł���p�ł���@p.881
- �M���b�v�����ɂ���āC�ߗזE�̓V�O�i���������L����@p.884
- ���ꂼ����זE�͓���̑g�ݍ��킹�̍זE�O�V�O�i�����q�ɉ�������悤�Ƀv���O��������Ă���@p.884
- �זE����ނ��قȂ�Γ����זE�O�V�O�i���ɑ��ĈقȂ鉞�������邱�Ƃ������@p.885
- �����r��̍זE�̉^���̓����t�H�Q���Z�x���z���̈ʒu�Ō��܂�@p.886
- �זE�����q�̎������Z���Ƃ��Ɍ���C�Z�x��v���ɕς��邱�Ƃ��ł���@p.886
- ��_�����f�͕W�I�זE���̓���̃^���p�N���̊����ڐ��䂵�ăV�O�i����`����@p.887
- �j����e�̂́C���K���h�ɂ���ĉe�������`�q���߃^���p�N�ł���@p.889
- �זE�\�ʎ�e�̃^���p�N�́C�C�I���`���l�������^�CG �^���p�N�����^�C�y�f�����^��3 ��ނɑ�ʂ����@p.891
- �����������זE�\�ʎ�e�̂̂قƂ�ǂ́C�����q��זE���V�O�i���^���p�N�̃l�b�g���[�N����ăV�O�i����`�B����@p.893
- �����̍זE���V�O�i���^���p�N�̓����_����GTP �̌����ɂ���Ċ���������C���q�X�C�b�`�Ƃ��ē����@p.895
- �זE���V�O�i���`�B�����͉̂����̑��x������C���ِ�������@p.897
- �זE���V�O�i���^���p�N�Ԃ̑��ݍ�p�́C���W���[���^���ݍ�p�h���C�����s���@p.897
- �זE�O�V�O�i���Z�x�̊ɂ₩�ȑ����ɑ��āC�זE�͕����̋@�\�𗘗p���ăI���E�I�t�̉������ł���@p.899
- �זE���V�O�i���`�B�l�b�g���[�N�́C�ʏ�C�t�B�[�h�o�b�N���[�v�𗘗p����@p.901
- �זE�̓V�O�i���ɑ��銴�x�߂ł���@p.902
- �܂Ƃ߁@p.903
���擪�s���E�I��\�ցA�͐߂̖ڎ��� �A�\�Q����
|
| 15��
�@�Q�� |
15-2. G�^���p�N�����^�זE�\�ʎ�e�́iGPCR�j�Ə����q�זE��������ɂ��V�O�i���`�B�i12���ځj�@ p.904
- GPCR ����̃V�O�i���𒆌p����O�ʑ�G �^���p�N�@p.905
- ��AMP �̐��Y�߂���G �^���p�N������@p.905
- ��AMP �̍�p�̂قƂ�ǂ́C��AMP �ˑ��̃^���p�N�L�i�[�[�iPKA�j������Ă���@p.908
- ������G �^���p�N�̓z�X�z���p�[�[C-b �̊������ɂ��C�C�m�V�g�[�����������̃V�O�i���`�B�o�H������������@p.909
- Ca2+ �͕��ՓI�ȍזE��������Ƃ��ē����@p.912
- Ca2+�Z�x�̎����I�ϓ��̕p�x���זE�̉����ɉe������@p.912
- �����זE�ł�Ca2+ �V�O�i���ւ̉����̑����́CCa2+/ �J�����W�������ˑ��^���p�N�L�i�[�[�iCaM�|�L�i�[�[�j������Ă���@p.914
- ������G �^���p�N�͒��ڃC�I���`���l���߂���@p.916
- �k�o�Ǝ��o�͊�k�N���I�`�h�ˑ��C�I���`���l���߂���GPCR �Ɉˑ�����@p.917
- �זE��������ƍy�f�A�������ɂ��זE�O�V�O�i�����啝�ɑ��������@p.919
- GPCR �̒E����͎�e�̂̃����_���Ɉˑ�����@p.920
- �܂Ƃ� 921
���擪�s���E�I��\�ցA�͐߂̖ڎ��� �A�\�Q����
|
| 15��
�@�R�� |
15-3. �y�f�����^�̍זE�\�ʎ�e�̂ɂ��V�O�i���`�B�i19���ځj�@ p.921
- ��e�̃`���V���L�i�[�[�͊���������Ǝ��g�������_������@p.922
- RTK ��̃����_���`���V���͍זE���V�O�i���^���p�N�Ƃ̐ڍ����ʂƂȂ�@p.923
- SH2 �h���C�������^���p�N���̓����_���`���V���c��Ɍ�������@p.924
- Ras �͒P�ʑ�GTP �A�[�[�̑傫�ȃX�[�p�[�t�@�~���[�ɑ�����@p.926
- RTK �̓A�_�v�^�[��GEF �����Ras ������������F�����r��̃V���E�W���E�o�G�̊Ⴉ�瓾��ꂽ�؋��@p.927
- Ras ��MAP �L�i�[�[�V�O�i���`�B���W���[��������������@p.928
- ����^���p�N�͕��s����MAP �L�i�[�[�iMAPK�j���W���[������������̂�h���@p.930
- Rho �t�@�~���[GTP �A�[�[�͍זE�\�ʎ�e�̂ƍזE���i���@�\�I�ɋ���������@p.931
- PI3�|�L�i�[�[�͍זE���Ɏ����̐ڒ����ʂ����@p.932
- PI3�|�L�i�[�[�|Akt �V�O�i���o�H�͓����זE���h�����Đ�������𑣐i����@p.934
- RTK ��GPCR ���d�Ȃ荇���ĉ����̃V�O�i���`�B�o�H������������@p.935
- �`���V���L�i�[�[��^��e�̂̊����́C�זE���̃`���V���L�i�[�[�ɂ��@p.935
- �T�C�g�J�C����e�̂�JAK–STAT �V�O�i���`�B�o�H�����������āC�V�O�i�����j�ɂ��₭���荞�ށ@p.937
- �^���p�N�`���V���z�X�t�@�^�[�[�ɂ���ă`���V���̃����_�������ɖ߂�@p.938
- TGFb �X�[�p�[�t�@�~���[�̃V�O�i���^���p�N�͎�e�̃Z����/ �g���I�j���L�i�[�[��Smad ����ē����@p.939
- �V�O�i���`�B�n��S�̂Ƃ��ė�������@p.941
- �ۂ̑������́C�q�X�`�W���L�i�[�[��^�̎�e�̂ɂ���Ċ����������2 �����n�V�O�i���`�B�o�H�Ɉˑ����Ă���@p.941
- ��e�̂̃��`�����ɂ��ۂ̑������͏�������@p.943
- �܂Ƃ߁@p.944
���擪�s���E�I��\�ցA�͐߂̖ڎ��� �A�\�Q����
|
| 15��
�@�S�� |
15-4. �V�O�i���`�B�o�H�ɂ�������ݓI��`�q���߃^���p�N�̕����̒����i5���ځj�@ p.946
- ��e�̃^���p�NNotch �͐��ݓI��`�q���߃^���p�N�ł���@p.946
- Wnt �^���p�N��Frizzled ��e�̂Ɍ�������b�|�J�e�j���̕�����j�Q����@p.948
- Hedgehog �^���p�N��Patched �ƌ������CSmoothened �̑j�Q����������@p.950
- ���d�X�g���X�h���Ɖ��ǗU���h����NFkB �ˑ��̃V�O�i���`�B�o�H�ɂ���ē����@p.952
- �܂Ƃ߁@p.954
���擪�s���E�I��\�ցA�͐߂̖ڎ��� �A�\�Q����
|
| 15��
�@�T�� |
15-5. �A���̃V�O�i���`�B�i6���ځj�@ p.955
- ���זE���ƍזE�ԏ��`�B�́C�����ƐA���œƗ��ɐi�����Ă����@p.955
- ��e�̃Z����/ �g���I�j���L�i�[�[�͐A���ł͍זE�\�ʎ�e�̂̍ł��傫�ȃO���[�v���\�����Ă���@p.956
- �G�`�����͊j�̓��ٓI�Ȉ�`�q���߃^���p�N�̕�����h���@p.957
- �I�[�L�V���A���̂̈ʒu���߂����ĐA���̐����p�^�[�������肷��@p.959
- �t�B�g�N�����͐ԐF�����C�N���v�g�N�����͐F�������m����@p.960
- �܂Ƃ߁@p.961
�͖����@p.962
���@���@p.964
���擪�s���E�I��\�ցA�͐߂̖ڎ��� �A�\�Q����
|
| 16�� |
�זE���i
�i�\���F4�� 53���ځj |
Ch.16 |
The Cytoskeleton |
| 16��
�@�P�� |
16-1. �זE���i�@�ۂ̎��ȏW���Ɠ��I�ȍ\���i15���ځj�@ p.965
- �זE���i�n�͓��I�œK����������@p.966
- �זE���i�͈���ȍ\��������@p.969
- �e��̍זE���i�@�ۂ́C��菬�����^���p�N�T�u���j�b�g����ł��Ă���@p.970
- �����̃v���g�t�B�������g����Ȃ�@�ۂɂ͗D�ꂽ����������@p.971
- �זE���i�̏d���̌`���ł́C�j�`���������i�K�ƂȂ�@p.973
- �`���[�u�����ƃA�N�`���͓��������ɕ���ŋɐ��̂���@�ۂ��`������@p.973
- �����ǂƃA�N�`���t�B�������g�͗��[�̍\�����قȂ��Ă��āC�L�����x���Ⴄ�@p.975
- �@�ۂ̃g���b�h�~����ԂƓ��I�s����́C�`���[�u�����ƃA�N�`���ɂ��k�N���I�`�h���������̌��ʐ�����@p.976
- �g���b�h�~����ԂƓ��I�s����͍זE���i�̐v���ȍč\����������@p.980
- �`���[�u�����ƃA�N�`���͐^�j�����̐i���ɂ����č��x�ɕۑ�����Ă����@p.982
- ���Ԍa�t�B�������g�̍\���́C���˂Ă˂�������荇�킹�R�C����ł���@p.983
- ���Ԍa�t�B�������g�͓����זE�ɕ����I�Ȉ��萫�������炷�@p.985
- ��܂��זE���i�@�ۂ̏d��������ω�������@p.987
- �ۂ̍זE�Ґ��ƍזE����͐^�j�����̍זE���i�̑����̂Ɉˑ����Ă���@p.989
- �܂Ƃ߁@p.991
���擪�s���E�I��\�ցA�͐߂̖ڎ��� �A�\�Q����
|
| 16��
�@�Q�� |
16-2. �זE���i�@�ۂ̒����i15���ځj�@ p.992
- �����ǂ̏d���j�`����g�|�`���[�u�������܂ރ^���p�N�����̂��s���@p.992
- �����זE�ł́C�����ǂ͒��S�̂���̂яo���@p.992
- �A�N�`���t�B�������g�̏d���j�`���͍זE���ŋN���邱�Ƃ������@p.996
- �d���j�`���̂����݂́C��K�͂ȃt�B�������g�̑g�D���ɉe������@p.998
- �@�ۂ̐L���͗V���T�u���j�b�g�Ɍ�������^���p�N���ɂ���Ē��߂����@p.999
- �A�N�`���t�B�������g����є����ǂ́C�ؒf�^���p�N�ɂ���Ē����Ɠ��I�ӂ�܂������䂳���@p.1000
- �@�ۂ̑��ʂɌ�������^���p�N���͑@�ۂ̈��艻��s���艻�ɂ������@p.1001
- �@�ۖ��[�ƍ�p����^���p�N���ő@�ۂ̓��Ԃ��啝�ɕς��@p.1002
- �ʎ�̃^���p�N�����}���ɐL����������ǖ��[�̐�����ς���@p.1003
- �זE���i�@�ۂ͍זE���̂�荂���̍\���̂ɑg�D�������@p.1005
- ���Ԍa�t�B�������g�͉ˋ�����C���˂��ď�v�Ȕz������@p.1005
- ���낢��Ȑ��������ˋ��^���p�N�̓����ŁC�A�N�`���t�B�������g���炳�܂��܂ȍ\�����ł���@p.1006
- �t�B���~���ƃX�y�N�g�����̓A�N�`���t�B�������g�̖Ԗڍ\�������@p.1008
- �זE���i�v�f�͑����̉ӏ��ōזE���Ɍ�������@p.1009
- �܂Ƃ߁@p.1010
���擪�s���E�I��\�ցA�͐߂̖ڎ��� �A�\�Q����
|
| 16��
�@�R�� |
16-3. ���q���[�^�[�i9���ځj�@ p.1010
- �A�N�`��������[�^�[�^���p�N�̓~�I�V���E�X�[�p�[�t�@�~���[�ɑ�����@p.1011
- �����ǃ��[�^�[�^���p�N�̓L�l�V���ƃ_�C�j����2 ��ނł���@p.1014
- �~�I�V���ƃL�l�V���̍\���ގ��́C�i���㋤�ʂ̋N�����l��������@p.1015
- ���[�^�[�^���p�N��ATP �����������\���ω��Ƌ��������ė͂�����@p.1016
- ���[�^�[�^���p�N�̑��x�_�I�����͍זE�̋@�\�ɓK�����Ă���@p.1020
- ���[�^�[�^���p�N�͖��ň͂܂ꂽ���튯�̍זE���A�����s���Ă���@p.1021
- �זE���i�͓����RNA ���q���Ǎ݉�������@p.1022
- �זE�̓��[�^�[�^���p�N�̋@�\�߂���@p.1023
- �܂Ƃ߁@p.1025
���擪�s���E�I��\�ցA�͐߂̖ڎ��� �A�\�Q����
|
| 16��
�@�S�� |
16-4. �זE���i�ƍזE�̂ӂ�܂��i14���ځj�@ p.1025
- II �^�~�I�V���ƃA�N�`���t�B�������g�̊���ɂ���ċؓ��̎��k���N����@p.1026
- �ؓ��̎��k�͍זE������Ca2+ �Z�x�̋}���ȏ㏸�ɂ���ċN����@p.1028
- �S�͌����ɐv���ꂽ�@�B�ł���@p.1031
- �@�тƕږт͔����ǂƃ_�C�j���łł����^���̂��߂̍\���ł���@p.1031
- �L�������a���̂̍\�z�ɂ́C���I�Ȕ����ǂƁC�����̃��[�^�[�^���p�N�̑��ݍ�p���K�v�ł���@p.1034
- ���낢��ȍזE���ő̂̊����͂����@p.1036
- �A�N�`���̏d�����זE����ˏo������@p.1037
- �ڒ��ƈ�������ɂ���čזE�͑O�i����@p.1040
- Rho�@p.�^���p�N�t�@�~���[�̒��Ԃ��A�N�`���זE���i�̑�K�͂ȍ\���ĕ҂������N���� 1041
- �זE�O�V�O�i����Rho �t�@�~���[��3 ��ނ̃^���p�N��������������@p.1043
- �O������̃V�O�i���ōזE�̈ړ����������܂�@p.1045
- �����ǂƃA�N�`���̍זE���i�ǂ������A�����čזE�S�̂̋ɐ���ړ��𐮍�������@p.1046
- �j���[�����̌`�Ԃ̕��G�ȓ��ꉻ�͍זE���i�ɂ���ċN����@p.1047
- �܂Ƃ߁@p.1050
�͖����@p.1050
���@���@p.1052
���擪�s���E�I��\�ցA�͐߂̖ڎ��� �A�\�Q����
|
| 17�� |
�זE�����@
�i�\���F8�� 64���ځj
|
Ch.17 |
The Cell Cycle |
| 17��
�@�P�� |
17-1. �זE�����̊T�v�i7���ځj�@ p.1054
- �^�j�����̍זE������4 �̎����ɕ������@p.1054
- ������^�j�����̍זE��������n�͎��Ă���@p.1056
- �y��̕ψى�͂���C�זE��������n����`�I�ɉ�͂��邱�Ƃ��ł���@p.1056
- ����������g���ƍזE��������n���w�I�ɉ�͂ł���@p.1057
- �M���ނ̔|�{�זE���g���āC�זE��������n�̌������ł���@p.1059
- �זE�����̐i�s������������@�͂��낢�날��@p.1059
- �܂Ƃ߁@p.1060
���擪�s���E�I��\�ցA�͐߂̖ڎ��� �A�\�Q����
|
| 17��
�@�Q�� |
17-2. �זE��������n�i7���ځj�@ p.1060
- �זE��������n�͍זE�����̎�v�ȉߒ��̊J�n���ƂȂ�@p.1060
- �זE�����̐���n�͎����I�Ɋ����������T�C�N�����ˑ��L�i�[�[�iCdk�j�ɂ���Ă���@p.1062
- �}���������_����Cdk �j�Q�^���p�N�iCKI�j��Cdk ������}������@p.1063
- �זE��������n�͎����I�ȃ^���p�N�����Ɉˑ�����@p.1064
- �זE�����̐���ɂ͓]�ʒ��߂��֗^����@p.1065
- �זE��������n�́C�����w�����̃X�C�b�`�̃I���E�I�t�Ő��藧���Ă���@p.1065
- �܂Ƃ߁@p.1067
���擪�s���E�I��\�ցA�͐߂̖ڎ��� �A�\�Q����
|
| 17��
�@�R�� |
17-3. S���i4���ځj�@ p.1067
- S-Cdk ��DNA �������J�n������͎̂���������1 ��ł���@p.1067
- ���F�̂̔{���ɂ̓N���}�`���\���̔{�����K�v�ł���@p.1069
- �R�q�[�V�����o�����F���̂�ڒ�����@p.1070
- �܂Ƃ߁@p.1071
���擪�s���E�I��\�ցA�͐߂̖ڎ��� �A�\�Q����
|
| 17��
�@�S�� |
17-4. �L�������i20���ځj�@ p.1071
- M-Cdk �͗L������̊J�n�𑣂��@p.1071
- �L������̏��߂ɁC�E�����_�ɂ��M-Cdk ������������@p.1074
- �R���f���V���͔{���������F�̂̕����ɕK�v�Ȋ��ݒ������@p.1075
- �L�������a���͔̂����ǂ��W�܂��Ăł���@p.1075
- �����Ljˑ����[�^�[�^���p�N���a���̂̌`���Ƌ@�\���i��@p.1077
- 2 �̋@�\���������ē�ɐ��L�������a���̂��`������@p.1077
- �זE�����̏��߂ɒ��S�̂��{������@p.1078
- M-Cdk �͑O���ɖa���̌`�����J�n������@p.1078
- �����זE�ɂ�����a���̌`���̊����ɂ͊j�����K�v�ł���@p.1079
- �L���������ɔ����ǂ̕s���肳���傫�������@p.1080
- ��������F�͓̂�ɖa���̂̌`���𑣂��@p.1081
- �����̂��o�����F���̂�a���̂ɕt��������@p.1082
- ��������͎��s����̎Y���ł���@p.1083
- ���F�͖̂a���̏�ł������̗͂ň��������ړ�����@p.1085
- APC/C ���o�����F���̕����ƗL�������𑣂��@p.1087
- �ɂƌ���Ă��Ȃ����F�͎̂o�����F���̂̕�����j�~���� �� �a���̌`���̃`�F�b�N�|�C���g�@p.1088
- ���F�̂͌��A �ƌ��B �ŕ�������@p.1089
- �����������F�̂͏I���ɖ��j�̒��ɔ[�܂�@p.1090
- ��������͗L�����B�ɓ��L�̊j����̌`�Ԃł���@p.1090
- �܂Ƃ߁@p.1092
���擪�s���E�I��\�ցA�͐߂̖ڎ��� �A�\�Q����
|
| 17��
�@�T�� |
17-5. �זE�������i9���ځj�@ p.1092
- ���k�̃A�N�`����II �^�~�I�V�����זE������̗͂����o���@p.1093
- RhoA �̋Ǐ��I�����������k�̍\�z�Ǝ��k�𑣂��@p.1094
- �L�������a���̂̔����ǂ������זE�̕����ʂ����肷��@p.1095
- �����A���ł͊u���`���̂��זE�������U������@p.1097
- �זE������ɂ͖��ŕ�܂ꂽ�זE���튯�����זE�ɕ��z����Ȃ���Ȃ�Ȃ��@p.1098
- �a���̂̈ʒu��ς��Ĕ�Ώ̕���������זE������@p.1099
- �זE������Ȃ��ŗL�����N���邱�Ƃ�����@p.1099
- G1 ���ɂ�Cdk ���s������Ԃɂ���@p.1100
- �܂Ƃ߁@p.1101
���擪�s���E�I��\�ցA�͐߂̖ڎ��� �A�\�Q����
|
| 17��
�@�U�� |
17-6. �זE�̕���Ɛ����̒����i3���ځj�@ p.1101
- �������i���q���זE����𑣐i����@p.1102
- ����Ȕ��Ԃɓ������זE�͕����x�点��@p.1103_
- �܂Ƃ߁@p.1010
���擪�s���E�I��\�ցA�͐߂̖ڎ��� �A�\�Q����
|
| 17��
�@�V�� |
17-7. ���q���[�^�[�i9���ځj�@�@ p.1010
- �A�N�`��������[�^�[�^���p�N�̓~�I�V���E�X�[�p�[�t�@�~���[�ɑ�����@p.1011
- �����ǃ��[�^�[�^���p�N�̓L�l�V���ƃ_�C�j����2 ��ނł���@p.1014
- �~�I�V���ƃL�l�V���̍\���ގ��́C�i���㋤�ʂ̋N�����l��������@p.1015
- ���[�^�[�^���p�N��ATP �����������\���ω��Ƌ��������ė͂�����@p.1016
- ���[�^�[�^���p�N�̑��x�_�I�����͍זE�̋@�\�ɓK�����Ă���@p.1020
- ���[�^�[�^���p�N�͖��ň͂܂ꂽ���튯�̍זE���A�����s���Ă���@p.1021
- �זE���i�͓����RNA ���q���Ǎ݉�������@p.1022
- �זE�̓��[�^�[�^���p�N�̋@�\�߂���@p.1023
- �܂Ƃ߁@p.1025
���擪�s���E�I��\�ցA�͐߂̖ڎ��� �A�\�Q����
|
| 17��
�@�W�� |
17-8. �זE���i�ƍזE�̂ӂ�܂��i14���ځj 1025
- II �^�~�I�V���ƃA�N�`���t�B�������g�̊���ɂ���ċؓ��̎��k���N���� p.1026
- �ؓ��̎��k�͍זE������Ca2+ �Z�x�̋}���ȏ㏸�ɂ���ċN����@p.1028
- �S�͌����ɐv���ꂽ�@�B�ł���@p.1031
- �@�тƕږт͔����ǂƃ_�C�j���łł����^���̂��߂̍\���ł���@p.1031
- �L�������a���̂̍\�z�ɂ́C���I�Ȕ����ǂƁC�����̃��[�^�[�^���p�N�̑��ݍ�p���K�v�ł���@p.1034
- ���낢��ȍזE���ő̂̊����͂����@p.1036
- �A�N�`���̏d�����זE����ˏo������@p.1037
- �ڒ��ƈ�������ɂ���čזE�͑O�i����@p.1040
- Rho �^���p�N�t�@�~���[�̒��Ԃ��A�N�`���זE���i�̑�K�͂ȍ\���ĕ҂������N�����@p.1041
- �זE�O�V�O�i����Rho �t�@�~���[��3 ��ނ̃^���p�N��������������@p.1043
- �O������̃V�O�i���ōזE�̈ړ����������܂�@p.1045
- �����ǂƃA�N�`���̍זE���i�ǂ������A�����čזE�S�̂̋ɐ���ړ��𐮍�������@p.1046
- �j���[�����̌`�Ԃ̕��G�ȓ��ꉻ�͍זE���i�ɂ���ċN����@p.1047
- �܂Ƃ߁@p.1050
�͖����@p.1050
���@���@p.1052
���擪�s���E�I��\�ցA�͐߂̖ڎ��� �A�\�Q����
|
| 18�� |
�A�|�g�[�V�X
�i�\���F1�� 9���ځj |
Ch.18 |
Apoptosis |
| 18��
�P�� |
- �v���O�����זE���͕s�v�ȍזE��r������@p.1115
- �A�|�g�[�V�X���N�������זE�͐����w�I�Ɏ��ʂł���@p.1117
- �A�|�g�[�V�X�́C�J�X�p�[�[�����זE���^���p�N�̘A�������Ɉˑ�����@p.1118
- �זE�\�ʂɂ���זE����e�̂��O���A�|�g�[�V�X�o�H������������@p.1120
- �����A�|�g�[�V�X�o�H�̓~�g�R���h���A�Ɉˑ�����@p.1121
- Bcl2 �^���p�N�������A�|�g�[�V�X�o�H�߂���@p.1121
- IAP �̓J�X�p�[�[��j�Q����@p.1124
- �זE�O�������q�͂��܂��܂Ȃ����݂ŃA�|�g�[�V�X��j�Q����@p.1126
- �A�|�g�[�V�X���ߏ肠�邢�͕s�\�����Ǝ����ɂȂ��邱�Ƃ�����@p.1127
- �܂Ƃ߁@p.1128
�͖����@p.1128
���@���@p.1129
���擪�s���E�I��\�ցA�͐߂̖ڎ��� �A�\�Q����
|
| ��X�� |
�זE�̂���Љ�
�i7�� 36�� 359���� 466�Łj
|
Part V. |
Cells in Their Social Context |
| 19�� |
�זE�����C�זE�ڒ��C
�זE�O�}�g���b�N�X�i�\���F7�� 68���ځj |
Ch.19 |
Cell Junctions, Cell Adhesion,
and the Extracellular Matrix |
| 19�́@
�P�� |
19-1. �J�h�w�����ƍזE�Ԑڒ��i15���ځj�@ p.1133
- �J�h�w�����͂����铮���ł�Ca2+ �ˑ��זE�Ԑڒ��Ɋ֗^����@p.1135
- �Ғœ����̃J�h�w�����E�X�[�p�[�t�@�~���[�͉��S��ނ��̃^���p�N������Ȃ�C�V�O�i���@�\�������̂������@p.1136
- �J�h�w�����͓���e�a���ɂ��ڒ����i��@p.1137
- ���ɂ����Ғœ����זE��I��I�ȍזE�Ԑڒ��ɂ���čďW�������g�D���Đ�������@p.1139
- �J�h�w�����͍זE�̑I��I�W���߂���@p.1140
- Twist �����|�ԏ[�D�J�ڂ߂���@p.1141
- �J�e�j���͌ÓT�I�J�h�w�������A�N�`���זE���i�ƘA��������@p.1142
- �ڒ������̓A�N�`���𗘗p���ėׂ荇���זE�ɋ����^�����N��������@p.1142
- �f�X���\�[�������ɂ���ď��͋��ՂɂȂ�@p.1143
- �זE�Ԑڒ��ōזE�����փV�O�i���𑗂�@p.1145
- �Z���N�`���͌����̒��ňꎞ�I�ȍזE�Ԑڒ���}���@p.1145
- �Ɖu�O���u�����E�X�[�p�[�t�@�~���[�ɑ�����^���p�N����Ca2+�Ɉˑ����Ȃ��זE�ڒ���}���@p.1146
- �V�i�v�X�`���ł͑���̍זE�ڒ����q�����s���ē����@p.1147
- ����^���p�N���ڑ������̂��`��������@p.1148
- �܂Ƃ߁@p.1149
���擪�s���E�I��\�ցA�͐߂̖ڎ��� �A�\�Q����
|
| 19��
�@�Q�� |
19-2. ���������Ə��̍\���i5���ځj�@ p.1150
- �����������זE�Ԍ����ӂ����C�_��������Ė���������@p.1150
- �ڑ������̂̑���^���p�N���זE���B�̐���ɕs���Ȗ�ڂ��ʂ����@p.1153
- �זE�Ԑڒ��Ɗ�ꖌ�ɂ���ď��̒���ɐ������܂�@p.1155
- ���ʓ��זE�ɐ��𐧌䂷��ʂ̃V�O�i���o�H�@p.1157
- �܂Ƃ߁@p.1158
���擪�s���E�I��\�ցA�͐߂̖ڎ��� �A�\�Q����
|
| 19��
�@�R�� |
19-3. �זE����זE�ւ̒ʘH�F�M���b�v�����ƌ��`���A���i11���ځj�@ p.1158
- �M���b�v�����ɂ���čזE�͓d�C�I�ɂ���ӓI�ɂ��������ꂽ��ԂɂȂ�@p.1158
- �M���b�v�����̃R�l�N�\���͖��ђʌ^�̃R�l�L�V���E�T�u���j�b�g6 ����Ȃ�@p.1159
- �M���b�v�����͑��l�ȋ@�\�����@p.1161
- �זE�̓M���b�v�����̓��ߐ��߂���@p.1161
- �A���ł́C�M���b�v�����Ɠ����@�\�����`���A�����S���@p.1162
- �܂Ƃ߁@p.1163
���擪�s���E�I��\�ցA�͐߂̖ڎ��� �A�\�Q����
|
| 19��
�@�S�� |
19-4. ��ꖌ�i5���ځj�@ p.1164
- ��ꖌ�͂��ׂĂ̏��̓y��ƂȂ�ƂƂ��ɁC�����̔���זE���@p.1164
- ���~�j���͊�ꖌ�̍ł��d�v�Ȑ����ł���@p.1165
- IV �^�R���[�Q��������Ɗ�ꖌ�ɒ��͑ϐ���������@p.1166
- ��ꖌ�͑��l�ȋ@�\�����@p.1167
- �܂Ƃ߁@p.1169
���擪�s���E�I��\�ցA�͐߂̖ڎ��� �A�\�Q�������擪�s���E�I��\�ցA�͐߂̖ڎ��� �A�\�Q����
|
| 19��
�@�T�� |
19-5. �C���e�O�����ƍזE�|�}�g���b�N�X�Ԑڒ��i8���ځj�@ p.1169
- �C���e�O�����͖��ђʂ̃w�e����ʑ̂ł���C�זE���i�ƘA������@p.1170
- �C���e�O�����͊����^�ƕs�����^�̃R���z���[�V�����ω�������@p.1170
- �C���e�O�����̌��ׂ͂��낢��Ȉ�`�a�̌����ƂȂ�@p.1172
- �C���e�O�����͏W���������ڒ����`������@p.1174
- �זE�O�}�g���b�N�X�Ԍ������C���e�O��������čזE�̑��B�Ɛ����𐧌�����@p.1175
- �C���e�O�����́C�זE�Ǝx���̂̐ڒ����ʂɍזE���V�O�i���^���p�N���W��������@p.1176
- �C���e�O�����͍זE���ɋǏ��I�ȍ��ق����o���@p.1177
- �܂Ƃ߁@p.1178
���擪�s���E�I��\�ցA�͐߂̖ڎ��� �A�\�Q����
|
| 19��
�@�U�� |
19-6. �����̌����g�D�̍זE�O�}�g���b�N�X�i18���ځj�@ p.1178
- �זE�O�}�g���b�N�X�̓}�g���b�N�X�̒��ɂ���זE�����C���������߂�@p.1179
- �O���R�T�~�m�O���J���iGAG�j���͑傫�ȋ�Ԃ��߁C���a�Q�����`������@p.1179
- �q�A�����i���͋�ԏ[�U�܂ł���Ɠ����ɁC�g�D�̌`�Ԍ`����C���̉ߒ��ɂ����čזE�̈ړ���������@p.1180
- �v���e�I�O���J����GAG �����R�A�^���p�N�ɋ��L�������Ăł������q�@p.1181
- �v���e�I�O���J���͕���^���p�N�̊����߂���@p.1182
- �זE�\�ʂ̃v���e�I�O���J���͕⏕��e�̂Ƃ��ē����@p.1183
- �זE�O�}�g���b�N�X�̎�v�ȃ^���p�N���Ƃ��ẴR���[�Q���ށ@p.1184
- �R���[�Q�����͖|���Ɉ�A�̏C������@p.1186
- �����Ƀv���y�v�`�h���v���R���[�Q�����q����菜���ꂽ��C���@�ۂ̌`�����n�܂�@p.1187
- ���傳�ꂽ���@�ی����R���[�Q���͌��@�ۂ�g�D������@p.1187
- �זE�̓}�g���b�N�X�ɂ���ڂ����͂ɂ���āC���債���R���[�Q�����@�ۂ̑g�D����������@p.1189
- �G���X�`���͑g�D�ɒe�͐���^����@p.1189
- �t�B�u���l�N�`���͍זE�O�^���p�N�Ń}�g���b�N�X�ւ̍זE�̕t����������@p.1191
- �זE�����������钣�͂��t�B�u���l�N�`�����@�ۂ̏W���߂���@p.1191
- �t�B�u���l�N�`����RGD ���`�[�t�ɂ���ăC���e�O�����Ɍ�������@p.1193
- �זE�̓}�g���b�N�X�̐��������łȂ����������Ȃ��Ă͂Ȃ�Ȃ��@p.1193
- �}�g���b�N�X�����͍זE�ߕӂŏW���I�ɋN����@p.1194
- �܂Ƃ߁@p.1195
���擪�s���E�I��\�ցA�͐߂̖ڎ��� �A�\�Q����
|
| 19��
�@�V�� |
19-7. �A���̍זE���i6���ځj�@ p.1195
- �זE�ǂ̑g���͍זE�̎�ނɂ��قȂ�@p.1195
- �זE�ǂ͒��͂ɋ����C�A���זE�ɂ͖c������������@p.1197
- �ꎟ�זE�ǂ̓Z�����[�X���@�ۂ��y�N�`�������ނ̖Ԗڍ\���ɕ҂ݍ��܂ꂽ�`�����Ă���@p.1197
- �זE�ǂ̒����̕������A���זE�̐����𐧌䂷��@p.1199
- �����ǂ��זE�ǒ����̌��������߂�@p.1200
- �܂Ƃ߁@p.1202
�͖����@p.1202
���@���@p.1204
���擪�s���E�I��\�ցA�͐߂̖ڎ��� �A�\�Q����
|
| 20�� |
�����i�\���F5�� 58���ځj�@ |
Ch.20 |
Cancer |
| 20��
�@�P�� |
20-1. ����Ɍ�������I�i���ߒ��i18���ځj�@ p.1205
- ����זE�͖������ɑ��B���C�ق��̑g�D�ɂ��݂��@p.1206
- �قƂ�ǂ̂����1 �ُ̈�זE���甭������@p.1207
- ����זE�ɂ͑̍זE�ψق��܂܂��@p.1208
- �P��̕ψق����ł́C����ɂƂ��ď\���łȂ��@p.1209
- ����͏������ُ킳�𑝂��זE���炵�����ɔ��B����@p.1210
- �q�{��͑��������ŗ\�h�ł���@p.1211
- ��ᇂ̓����_���ȕψق̌p���Ǝ��R�I�����J��Ԃ��Ȃ���i�s����@p.1212
- ����זE�ɒ~�ς���G�s�W�F�l�e�B�b�N�ȕω��ɂ̓N���}�`���\����DNA �̃��`�����̌p��������@p.1213
- �q�g�̂���זE�͈�`�I�ɕs����ł���@p.1214
- �זE����זE�����̐���Ɍ��ׂ�����Ƃ���ɂȂ�₷���@p.1215
- ����זE�͕��ʁCDNA �����₻�̑��̃X�g���X����ƕω�����@p.1216
- �q�g�̂���זE�͑��B�̐����@�\�����蔲����@p.1217
- �����̎�ᇂ̈ێ��ɏ��W�c�̂��זE�������Ă���@p.1217
- ���זE�������邵���݁@p.1218
- �]�ڂɂ́C��������זE���َ��̊��Ő��������B����K�v������@p.1220
- ��ᇂ͌��ǐV����U������@p.1220
- ��ᇂ̔���������̔����ɉe������@p.1222
- ����̑��B�ɂ�������Ă��鑽�l�Ȑ����@p.1223
- �܂Ƃ߁@p.1223
���擪�s���E�I��\�ցA�͐߂̖ڎ��� �A�\�Q����
|
| 20��
�@�Q�� |
20-2. �\�h�ł��锭����̌����i5���ځj�@ p.1224
- �S���ł͂Ȃ����C�����̔�����v����DNA �ɑ�����^����@p.1225
- ������C�j�V�G�[�^�[��DNA �����邪�C������v�����[�^�[�͑������Ȃ��@p.1226
- �E�C���X�Ȃǂ̊������q�g�̂���̌����ɂȂ邱�Ƃ�����@p.1227
- ��������q�肷��C��������������@��������@p.1229
- �܂Ƃ߁@p.1230
���擪�s���E�I��\�ցA�͐߂̖ڎ��� �A�\�Q����
|
| 20��
�@�R�� |
20-3. ���ɏd�v�Ȉ�`�q�̒T���i9���ځj�@ p.1230
- �@�\�l���ψقƋ@�\�����ψق͓���@���قȂ�@p.1231
- ���g���E�C���X�͍זE�̋�����ς��邪���`�q���^�э��ނ��Ƃ�����@p.1232
- ���܂��܂Ȃ����`�q�̒T������1 �̈�`�q�CRas ���˂��~�߂�ꂽ�@p.1233
- �܂�Ȉ�`������̌����ɂ��C����}����`�q�����肳�ꂽ�@p.1234
- ����}����`�q�͎�ᇂ��̂��̂̌������������ł���@p.1235
- ����}����`�q�́C��`�q�̕ω��ł��G�s�W�F�l�e�B�b�N�Ȃ����݂ł��s����������@p.1235
- ����ŕψق��N��������`�q�͂��낢��Ȃ����݂ʼnߏ芈���^�ɂȂ�@p.1237
- ���ɏd�v�Ȉ�`�q�̒T���͑����Ă���@p.1239
- �܂Ƃ߁@p.1240
���擪�s���E�I��\�ցA�͐߂̖ڎ��� �A�\�Q����
|
| 20��
�@�S�� |
20-4. ����זE�̋����Ƃ��̕��q���x���ł̉��i13���ځj�@�@ p.1241
- ���ƈ�`�q���σ}�E�X�̌������C���ɏd�v�Ȉ�`�q�̋@�\�𖾂ɖ𗧂��Ă����@p.1241
- ���ɏd�v�Ȉ�`�q�̑����͍זE���B�߂��Ă���@p.1242
- ����זE�ōזE�����̐i�s�ƍזE�̐����̒��߂����킹��o�H�͕ʂ�������Ȃ��@p.1244
- �A�|�g�[�V�X�߂����`�q�̕ψق́C����זE�����ׂ��łȂ��Ƃ��ɑ���������@p.1245
- p53 ��`�q�̕ψقɂ���āCDNA ���������Ă����������B���邪��זE�������@p.1246
- DNA ��ᇃE�C���X�́C���ƂȂ邪��}���^���p�N�̍�p��j�~����@p.1247
- �]�ڂɂȂ����ᇍזE�̕ψق͂܂���ɕ�܂�Ă���@p.1249
- �咰����͖ڂɌ�����ω��𑱂��Ȃ���������i������@p.1250
- �����̏d�v�Ȉ�`�q�̑������啔���̑咰����ɋ��ʂ��Ă݂���@p.1251
- DNA �̕s���C���n�Ɍ���������咰���������@p.1254
- ��ᇂ̒i�K�I�i�s�͓���̕ψقƊW�Â����邱�Ƃ������@p.1254
- ����͂��ꂼ��ɋN������`�q�����̏����f���Ă���@p.1256
- �܂Ƃ߁@p.1256
���擪�s���E�I��\�ցA�͐߂̖ڎ��� �A�\�Q����
|
| 20��
�@�T�� |
20-5. ���Â̌���Ə����i12���ځj�@ p.1256
- ���Â̒T���͂ނ����������C��]�͂���@p.1257
- �]���̎��Ö@�́C����זE����`�I�ɕs����ł���זE�����̃`�F�b�N�|�C���g�������������Ă��邱�Ƃ𗘗p���Ă���@p.1257
- ��ᇂ̈�`�I�s���萫�̓����𗘗p����V��@p.1257
- ��`�I�ɕs����Ȃ��߁C����͒i�K�I�Ɏ��Âɒ�R���ɂȂ�@p.1259
- ����̐����w�̒m������V�������Ö@�����܂����@p.1260
- ����̔�����^���p�N��j�Q���鏬���q��v�ł���@p.1260
- ��ᇂ̌��ǂ͂��Â̗��ɂ��Ȃ����W�I�ł���@p.1262
- ����̎�ᇂɑ���Ɖu���������Ď��Âł��邪����������낤�@p.1262
- �����̖�̓������^�����ÂɗL����������Ȃ��@p.1263
- ��`�q�����v���t�@�C����͂ŗՏ��I�ɖ𗧂���̕��ނ��ł���@p.1264
- �ۑ�͈ˑR�Ƃ��ĎR�ς��Ă���@p.1264
- �܂Ƃ߁@p.1265
�͖����@p.1265
���@���@p.1267
���擪�s���E�I��\�ցA�͐߂̖ڎ��� �A�\�Q����
|
| 21�� |
�L�����B�F��������C���B�זE�C���i�\���F6�� 36���ځj |
Ch.21 |
Sexual Reproduction : Meiosis, Germ Cells and Fertilization |
| 21��
�@�P�� |
21-1. �L�����B�Ƃ��i4���ځj�@ p.1269
- �����Ȑ^�j�����ł́C��{�̂̊��Ԃ͔��ɒZ���@p.1269
- ��������`�I���l���ށ@p.1271
- �L�����B�͐����̂ɗL���ł���@p.1271
- �܂Ƃ߁@p.1272
���擪�s���E�I��\�ցA�͐߂̖ڎ��� �A�\�Q����
|
| 21��
�@�Q�� |
21-2. ���������i9���ځj�@ p.1272
- �z��q��2 ��̓��ʂȍזE����Ő�����@p.1272
- ���������������F�́i�Ɛ����F�́j�͌�������̑�ꕪ��̏��߂ɑ�����@p.1274
- �������F�̑��ŃV�i�v�g�l�}�\�����`�������@p.1275
- �����������ٓI�����̌����^���p�N���C�������F�̕�����S���@p.1276
- ��������ł͂悭�ԈႢ���N����@p.1278
- ��������`�q�̍ĕҐ��𑣐i����@p.1279
- �����͌����ɐ��䂳��Ă���@p.1280
- �M���ނ̗Y�Ǝ��ł͌�������̐���̂��������قȂ�@p.1280
- �܂Ƃ߁@p.1281
���擪�s���E�I��\�ցA�͐߂̖ڎ��� �A�\�Q����
|
| 21��
�@�R�� |
21-3. �M���ނ̎n�����B�זE�Ɛ������i5���ځj�@�@ p.1282
- �M���ނ���ł͎��͂���̃V�O�i���Ŏn�����B�זE�����߂���@p.1282
- �n�����B�זE�͔����r��̐��B�B�Ɉړ�����@p.1283
- Sry ��`�q���M���ނ̔����r��̐��B�B���ɕ�������悤�w�߂���@p.1283
- �L�����B�͓����̎�ނɂ���đ傫���قȂ�@p.1285
- �܂Ƃ߁@p.1286
���擪�s���E�I��\�ցA�͐߂̖ڎ��� �A�\�Q����
|
| 21��
�@�S�� |
21-4. ���i5���ځj�@ p.1287
- ���͂��ꂾ���Ŕ����ł���悤���x�ɐ�剻���Ă���@p.1287
- ���̌`���͒i�K��ǂ��Đi�s����@p.1288
- ����זE�͓��ʂȋ@�\�ɂ���đ傫����������@p.1290
- �q�g�̗���זE�͂قƂ�ǂ����n�����Ɏ��ʁ@p.1291
- �܂Ƃ߁@p.1292
���擪�s���E�I��\�ցA�͐߂̖ڎ��� �A�\�Q����
|
| 21��
�@�T�� |
21-5. ���@�q�i4���ځj�@ p.1292
- ���q�͎��g��DNA �𗑂ɓn���Ƃ����ړI�ɓK���������Ă���@p.1292
- �M���ނ̐����͂��������q�Y���Ă���@p.1293
- ���q�͑��j�זE�Ƃ��Ĕ�������@p.1294
- �܂Ƃ߁@p.1296
���擪�s���E�I��\�ցA�͐߂̖ڎ��� �A�\�Q����
|
| 21��
�@�U�� |
21-6. ��@���i9���ځj�@ p.1297
- �ː����ꂽ���q�͎��̐��B�ǂŎ\�͂�@p.1297
- �\���l���������q�͓����тɌ������C��̔������N����@p.1298
- ���q�Ɨ��̗Z���̂����݂͂܂��悭�킩��Ȃ��@p.1298
- ���́C���q�Ƃ̗Z���ɂ��זE����Ca2+ �����Ŋ���������@p.1299
- ���̔�w�i�\�w�j�����ɂ��1 �����̐��q�����邱�ƂɂȂ�@p.1300
- ���q�̓Q�m�������ł͂Ȃ����S���̂��ڍ��q�ɂ������ށ@p.1301
- �q�g�̕s�D���Âɕϊv�������炵���̊O�Ɖ���B��Á@p.1301
- �܂Ƃ߁@p.1303
- ���@���@p.1304
���擪�s���E�I��\�ցA�͐߂̖ڎ��� �A�\�Q����
|
| 22�� |
���זE�����ɂ����锭���@
�i�\���F2�� 31���ځj |
Ch.22 |
Development of Multicellular Organisms |
| 22��
�@�P�� |
22-1. �����̔����̊�{�i18���ځj�@ p.1305
- �����̊�{�I�ȉ�U�w�I�����͋��ʂ��Ă���@p.1307
- ���זE�����ł͍זE�ԑ��ݍ�p���`�q���߂Ɋ֗^����^���p�N�����L�x�ł���@p.1308
- ����DNA �������̃v���O���������߂�@p.1309
- ��̑���ɂ��זE�Ԃ̑��ݍ�p�����ׂ���@p.1310
- �ψٓ����̌����ɂ�蔭���̉ߒ��𐧌䂷���`�q�����肳���@p.1311
- �ώ@�ł���ω������͂邩�ȑO�ɁC�זE�͔����ɂ�����錈����s���@p.1311
- �זE�͑̂̒��̈ʒu�f����ʒu�����L�����Ă���@p.1312
- �U���I�V�O�i���́C�ŏ��͓��ꂾ�����זE�Ԃɒ����������Ⴂ�����o���@p.1313
- ��Ώ̓I�ȍזE����ɂ��قȂ�o���זE��������@p.1313
- ���̃t�B�[�h�o�b�N�ɂ���Ώ̐����V���ɍ��o�����@p.1314
- ���̃t�B�[�h�o�b�N�Ńp�^�[�����ł��C�S�������̌��ʂ����o���āC�L���������炷�@p.1315
- �����̃V�O�i���o�H���J��Ԃ��g���p�^�[���`���𐧌䂷��@p.1316
- �����t�H�Q���͒������ɓ����U�����q�ŁC�Z�x���z���ʂ������@p.1316
- �V�O�i�����q�ɑ���זE�O�j�Q���q�ɂ��C�U�����q�ւ̉������ς��@p.1317
- �����V�O�i���͕����̈قȂ�����őg�D�ɍL����@p.1318
- �זE�ŗL�̃v���O�����ɂ�蔭���̎����\����߂���@p.1319
- �ŏ��̃p�^�[���͏����ȍזE�̈�Ŋm������C��̐����ɔ����i�K�I�ȗU���ɂ���Ė��m�������@p.1319
- �܂Ƃ߁@p.1320
���擪�s���E�I��\�ցA�͐߂̖ڎ��� �A�\�Q����
|
| 22��
�@�Q�� |
22-2. ����C. elegans - �X�̍זE���猩�锭���i13���ځj�@ p.1321
- ����C. elegans �͉�U�w�I�ɒP���ł���@p.1321
- �����r��̐����̍זE�̉^���͂قڊ��S�ɗ\���ł���@p.1322
- �ꐫ���ʈ�`�q�̎Y�������̔�Ώ̂ȕ�������߂�@p.1323
- �זE�ԑ��ݍ�p�ɂ��C�p�^�[���͒i�K�I�ɕ��G���𑝂��Ă����@p.1324
- ������p�ƈ�`�w�ɂ�蔭���̐���̂����ׁ݂C��`�q�N���[�j���O�Ɣz��ŕ��q�@�\�𖾂炩�ɂ���@p.1325
- �זE�͎��Ԍo�߂ƂƂ��ɔ����V�O�i���ւ̉�����ς���@p.1325
- �َ�����`�q�͔����̃^�C�~���O�𐧌䂷��@p.1326
- ����̎�ᇂɑ���Ɖu���������Ď��Âł��邪����������낤�@p.1262
- �����̖�̓������^�����ÂɗL����������Ȃ��@p.1263
- ��`�q�����v���t�@�C����͂ŗՏ��I�ɖ𗧂���̕��ނ��ł���@p.1264
- �ۑ�͈ˑR�Ƃ��ĎR�ς��Ă���@p.1264
- �܂Ƃ߁@p.1265
�͖����@p.1265
���@���@p.1267
���擪�s���E�I��\�ցA�͐߂̖ڎ��� �A�\�Q����
|
| 23�� |
��剻�����g�D�C���זE�Ƒg�D�̍Đ��@�i�\���F8�� 68���ځj |
Ch.23 |
Specialized Tissues, Stem Cells and Tissue |
| 23��
�@�P�� |
23-1. �\��Ɗ��זE�ɂ�邻�̍X�V�i11���ځj�@ p.1417
- �\��זE�͑��w����Ȃ�h���ǂ�����Ă���@p.1419
- �����r��̕\��זE�́C���n�ƂƂ��Ɋe��̈�`�q������ǂ��Ĕ�������@p.1420
- ���זE�w�̊��זE�͕\����X�V����@p.1420
- ���זE��2 �̖��זE���ʎ�̍זE�ɂȂ�Ƃ͌���Ȃ��@p.1421
- ���w�ɂ͊��זE�̂ق��ꎞ�I�ɑ��������זE���܂܂��@p.1422
- �ꎞ�I��������͐�������헪�̈ꕔ�ł���@p.1423
- ���זE���I��I�ɂ��Ƃ�DNA �����ێ�����g�D������@p.1424
- �ً}�ɐV�����זE���K�v�ɂȂ�Ɗ��זE�̕������x�͌��I�ɑ�������@p.1425
- �����̃V�O�i���̑��ݍ�p���\��̍X�V���x�z���Ă���@p.1426
- ���B�͔����Ƒމ����J��Ԃ��@p.1426
- �܂Ƃ߁@p.1428
���擪�s���E�I��\�ցA�͐߂̖ڎ��� �A�\�Q����
|
| 23��
�@�Q�� |
23-2. ���o����i4���ځj�@ p.1429
- �k�o�j���[�����͂˂ɒu��������Ă���@p.1429
- ���o�L�эזE�͈ꐶ�C���������� 1430
- �ꐶ�ێ������זE�̑啔���ŁC���i�̍X�V���݂��� �� �Ԗ��̌���e��זE�@p.1432
- �܂Ƃ߁@p.1433
���擪�s���E�I��\�ցA�͐߂̖ڎ��� �A�\�Q����
|
| 23��
�@�R�� |
23-3. �C���ƒ����i11���ځj�@ p.1434
- �x�̔x�E�ł́C�قȂ邪�ׂ荇���זE�����͂������@p.1434
- �C���̐��|��Ƃ͔t�זE�C�@�эזE�C�}�N���t�@�[�W�������ōs���Ă���@p.1434
- �����̓��ǂ͂ق��̂ǂ̑g�D���������X�V���Ă���@p.1436
- Wnt �V�O�i�������NJ��זE�����ێ�����@p.1438
- Notch �V�O�i���͒��זE�̑��l���𐧌䂷��@p.1439
- �G�t�����|Eph �V�O�i���͒����זE�̈ړ��𐧌䂷��@p.1440
- Wnt�CHedgehog�CPDGF�C�����BMP �V�O�i���o�H�̑g�ݍ��킹�͊��זE�̐��������߂�@p.1441
- �̑��͏����ǂƌ��t�̒��p���Ƃ��ē����Ă���@p.1442
- �̍זE��������Ƃ��̑��B�����i����@p.1443
- �g�D�̍X�V�͕K���������זE�Ɉˑ����Ă��Ȃ��F�X���̃C���X��������זE�̗�@p.1444
- �܂Ƃ߁@p.1445
���擪�s���E�I��\�ցA�͐߂̖ڎ��� �A�\�Q����
|
| 23��
�@�S�� |
23-4. ���ǁC�����p�ǂƓ���זE�i6���ځj�@ p.1445
- ���Ǖǂƃ����p�Ǖǂ̓����͂��ׂē���זE�������Ă���@p.1445
- �����[�זE�����ǐV���̐�삯�ƂȂ�@p.1446
- �Ⴄ��ނ̌��ǂ��`���������זE�͎�ނ��Ⴄ�@p.1447
- ������K�v�Ƃ���g�D��VEGF ����o���C����זE�Ԃ�Notch �V�O�i���o�H��VEGF �ւ̉����𐧌䂷��@p.1448
- ����זE����̃V�O�i���͎���זE�ƕ����؍זE�������Ǖǂ��`������@p.1450
- �܂Ƃ߁@p.1450
���擪�s���E�I��\�ցA�͐߂̖ڎ��� �A�\�Q����
|
| 23��
�@�T�� |
23-5. ���\�����זE�ɂ��X�V �� ���t�זE�̌`���i13���ځj�@ p.1450
- ���������������C�P���C�����p����3 ��ނɕ��ނ����@p.1451
- �������ł̌��t�זE�̐��Y�͎�ނ��Ƃɒ��߂���Ă���@p.1453
- �����ɂ͑������זE������@p.1454
- ���t�זE�͂��ׂđ��\�����זE���琶����@p.1456
- �����̕����Â��͒i�K��ǂ��Đi�s����@p.1456
- �����������Â����ꂽ�O��זE�����C��剻���������̐��𑝂₷�@p.1457
- ���זE�̓X�g���[�}�זE����̐ڐG�V�O�i���Ɉˑ����Ă���@p.1458
- �����߂�����q�͔|�{�זE�n�Œ��ׂ���@p.1459
- �Ԍ����̐����̓G���g���|�G�`���Ɉˑ�����@p.1459
- �D�����ƃ}�N���t�@�[�W�̐��Y�ɂ͕�����CSF ���e������@p.1460
- �����זE�̂ӂ�܂��ɂ͋��R�Ɍ��܂�v�f������@p.1461
- �זE�̐����̐���͑��B�̐���Ɠ������炢�d�v�ł���@p.1462
- �܂Ƃ߁@p.1462
���擪�s���E�I��\�ցA�͐߂̖ڎ��� �A�\�Q����
|
| 23��
�@�U�� |
23-6. ���i�̔����C�@�\���߁C�Đ��i5���ځj�@ p.1463
- �V�������i�ؑ@�ۂ͋؉�זE���Z�����Ăł���@p.1464
- �؍זE�̓^���p�N���̃C�\�^���ւ��Ď��g�̓�����ς�����@p.1465
- ���i�ؑ@�ۂ̓~�I�X�^�`���債�Ď��g�̐����𐧌�����@p.1465
- �ꕔ�̋؉�זE�́C�������Ȃ����זE�Ƃ��Đ��̒��ɂ����݂��Ă���@p.1466
- �܂Ƃ߁@p.1467
���擪�s���E�I��\�ցA�͐߂̖ڎ��� �A�\�Q����
|
| 23��
�@�V�� |
23-7. �@�ۉ�זE�Ƃ��̌`���̕ω� �� �����g�D�i9���ځj�@ p.1467
- �@�ۉ�זE�͉��w�V�O�i���ɉ������Đ�����ς���@p.1467
- �זE�O�}�g���b�N�X�͍זE�̌`��ڒ����ɉe����^���Č����g�D�זE�̕����Ɋ֗^����炵���@p.1468
- ����זE�͍��}�g���b�N�X�����@p.1469
- �قƂ�ǂ̍��͓�łł������`�̎��͂ɍ����@p.1470
- ���͓����̍זE�ɂ���Ă�������������Ă���@p.1472
- �j���זE�͍���זE����̃V�O�i���ɂ�萧�䂳��Ă���@p.1473
- ���b�זE�͑@�ۉ�זE���琶����@p.1474
- ���b�זE�����傷�郌�v�`���́C�ېH��}������t�B�[�h�o�b�N�@�\�ƂȂ�@p.1475
- �܂Ƃ߁@p.1476
���擪�s���E�I��\�ցA�͐߂̖ڎ��� �A�\�Q����
|
| 23��
�@�W�� |
23-8. ���זE�H�w�i10���ځj�@ p.1477
- �������זE�𗘗p����ƕa�C�̌��t�זE�����N�ȍזE�ɒu�������邱�Ƃ��ł���@p.1477
- �\��̊��זE�W�c�͔|�{�ɂ��g�債�đg�D�C���Ɏg����@p.1477
- �_�o���זE�͔|�{�ɂ�葀��ł���@p.1478
- �_�o���זE�͒����_�o�n�ɒ蒅�����B�ł���@p.1478
- ���̂̊��זE�͑g�D���ٓI�ł���@p.1479
- ES �זE�͑̂̂ǂ̕����ł����o�����Ƃ��ł���@p.1480
- ���ғ��ٓI��ES �זE�͖Ɖu����̖�����������@p.1481
- ES �זE�͑n��ƕa�C�̉�͂ɖ𗧂@p.1482
- �܂Ƃ߁@p.1482
- ���@���@p.1483
���擪�s���E�I��\�ցA�͐߂̖ڎ��� �A�\�Q����
|
| 24�� |
�a���́C�����C���R�Ɖu
�i�\���F3�� 36���ځj |
Ch.24 |
Pathogens, Infection, and Innate Immunity |
| 24��
�@�P�� |
24-1. �a���̂̊T���i9���ځj�@ p.1486
- �a���̂͏h��ɂƂ�����߂̓��ʂȂ����݂�i�������Ă����@p.1486
- �����̒����Ǐ�ɂ́C�a���̂ɂ����̂Əh��̔����ɂ����̂Ƃ�����@p.1487
- �a���̂͌n�������I�ɑ��l�ł���@p.1488
- �a���ۂ͓���̓Ő���`�q�������Ă���@p.1489
- �ۗނ���������͈قȂ�`����Ƃ镡�G�Ȑ��������@p.1494
- �E�C���X�͑��B�̂�����ʂŏh��זE�̑��u�Ɉˑ�����@p.1496
- �v���I���͊������^���p�N�ł���@p.1498
- �����ǂ��N�����a���̂́C����C�S���a���̑��̖��������Ɋ֘A������@p.1499
���擪�s���E�I��\�ցA�͐߂̖ڎ��� �A�\�Q����
|
| 24��
�@�Q�� |
24-2. �����̍זE�����w�i16���ځj�@ p.1501
- �a���͖̂h���ǂ����z���ďh��ɓ��荞�ݏZ�݂��@p.1501
- ���ɏZ�݂��a���̂͏h��̔r���@�\��������Ȃ���Ȃ�Ȃ��@p.1502
- �זE���̕a���̂́C�h��זE�ւ̐N���ƒE�o�̗����̂����݂�����Ă���@p.1504
- �E�C���X���q�͏h��זE�̕\�ʂɒ��ꂽ���q�Ɍ�������@p.1505
- �r���I���́C���Z���⏬�E�`���C���̔j��ɂ���ďh��זE�ɐN������@p.1506
- �ۂ͐H��p�𗘗p���ďh��זE�ɐN������@p.1507
- �זE���Ɋ���^�j�����́C�h��זE�ɐϋɓI�ɐN������@p.1508
- �����̕a���̂́C�h��זE�̖��ړ���ω�������@p.1511
- �E�C���X��ۂ́C�h��זE�̍זE���i�𗘗p���čזE�����ړ�����@p.1514
- �E�C���X�����͏h��זE�̑�ӂ�������@p.1517
- �a���̂͏h�吶���̍s����ω������ē`�d�𑣐i����@p.1518
- �a���̂͋}���ɐi������@p.1518
- �a���̂̍R���ψق́C�����̋@�\�ŋN����@p.1519
- ���̑����������E�C���X�̐i�����x�z����@p.1520
- �a���̖̂�ܑϐ����͂܂��܂��[���ɂȂ��Ă���@p.1521
- �܂Ƃ߁@p.1524
���擪�s���E�I��\�ցA�͐߂̖ڎ��� �A�\�Q����
|
| 24��
�@�R�� |
24-3. ������W�����ǂƎ��R�Ɖu�n�i11���ځj�@ p.1524
- ���\�ʂƃf�t�F���V���͊����̗\�h�ɖ𗧂@p.1525
- �q�g�̍זE�͕a���̂ɕۑ�����Ă�����������ʂ���@p.1526
- ��̂̊������ɂ��C�a���̂�H��p��n���̕W�I�ɂ���@p.1528
- Toll �l�^���p�N��NOD �^���p�N�͌Â����炠��p�^�[�����ʎ�e�̃t�@�~���[�ł���@p.1530
- �H�זE�͕a���̂�T���o���C���ݍ���Ŕj��@p.1531
- �����������}�N���t�@�[�W���������ʂ̉��ǔ����Ɋ֗^����@p.1533
- �E�C���X�����זE�͎v��������i�ŃE�C���X�̑��B��h���@p.1534
- �i�`�������L���[�זE�̓E�C���X�����זE�����E������@p.1535
- ����זE�͎��R�Ɖu�n�ƓK���Ɖu�n���Ȃ�����������@p.1536
- �܂Ƃ߁@p.1537
- ���@���@p.1537
���擪�s���E�I��\�ցA�͐߂̖ڎ��� �A�\�Q����
|
| 25�� |
�K���Ɖu�i�\���F5�� 52���ځj |
Ch.25 |
The Adaptive Immune System |
| 25��
�@�P�� |
25-1. �����p���ƓK���Ɖu�̍זE�w�i9���ځj�@ p.1540
- �K���Ɖu�ɂ̓����p�����K�v�ł���@p.1540
- ���R�Ɖu�n�ƓK���Ɖu�n�͘A�g���ē����@p.1541
- B �זE�͍����ō���CT �זE�͋��B�ō����@p.1543
- �K���Ɖu�n�̓N���[���I���ɂ���ē����@p.1544
- �قƂ�ǂ̍R���͕����̃����p���N���[��������������@p.1545
- �Ɖu�L���ɂ́C�N���[���̑��B�ƃ����p���̕������֗^����@p.1545
- �Ɖu���e�̂������Ŏ��ȍR���͒ʏ�C�U������Ȃ��@p.1547
- �����p���͖��������p�튯���������z���Ă���@p.1549
- �܂Ƃ߁@p.1551
���擪�s���E�I��\�ցA�͐߂̖ڎ��� �A�\�Q����
|
| 25��
�@�Q�� |
25-2. B�זE�ƍR���i9���ځj�@ p.1552
- B �זE�����R�̂ɂ́C�זE�\�ʂ̍R����e�̂ƕ���^���p�N�Ƃ�����@p.1552
- �T�^�I�ȍR�̂ɂ́C����̍R���������ʂ�2 ����@p.1552
- �R�̕��q��H ����L ������ł��Ă���@p.1552
- �R��H ���ɂ�5 �̃N���X������C�e�N���X�͈قȂ鐶���w�I���������@p.1553
- �R�̂ƍR���̑��ݍ�p�̋����́C�R���������ʂ̐��Ɛe�a���Ɉˑ�����@p.1557
- �R�̂�L ����H ���͒��̈�Ɖϗ̈悩��ł��Ă���@p.1558
- L ����H ���͔�������Ig �h���C������\������Ă���@p.1559
- �R���������ʂ͒��σ��[�v�ō\�������@p.1560
- �܂Ƃ߁@p.1561
���擪�s���E�I��\�ցA�͐߂̖ڎ��� �A�\�Q����
|
| 25��
�@�R�� |
25-3. �R�̂̑��l���ݏo���������i6���ځj�@ p.1562
- �R�̈�`�q��B �זE�̕����ɔ����ĕʁX�̈�`�q�f�Ђ���g�ݗ��Ă���@p.1562
- ��`�q�f�Ђ̕s���m�ȘA����V �̈�̑��l���傳����@p.1564
- B �זE���P��̓��ِ��������̂́CV(D)J �g���������䂳��邽�߂ł���@p.1565
- �R���͑̍זE���ψق�U�����čR�̉��������������@p.1566
- B �זE�͐��Y����R�̂̃N���X���ւ���@p.1567
- �܂Ƃ߁@p.1568
���擪�s���E�I��\�ցA�͐߂̖ڎ��� �A�\�Q����
|
| 25��
�@�S�� |
25-4. T�זE��MHC�^���p�N�i18���ځj�@ p.1569
- T �זE��e�́iTCR�j�͍R�̂Ɏ����w�e����ʑ̂ł���@p.1570
- T �זE�͎���זE�̍R���ɂ���Ċ������܂��͊��e������@p.1571
- �G�t�F�N�^�[�זE���Q��T �זE�́C���������W�I�זE�̎��E��U������@p.1572
- �G�t�F�N�^�[�w���p�[T �זE�͂ق��̎��R�Ɖu�n��K���Ɖu�n�̍זE�̊�������������@p.1573
- ���ߐ�T �זE�͂ق���T �זE�̊�����}������@p.1574
- T �זE��MHC �^���p�N�Ɍ��������ȃy�v�`�h�����ʂ���@p.1575
- MHC �^���p�N�́C�@�\���킩��O�ɈڐA�����œ��肳�ꂽ�@p.1575
- �N���XI �ƃN���XII ��MHC �^���p�N�́C�悭�����\���̃w�e����ʑ̂ł���@p.1576
- MHC �^���p�N�̓y�v�`�h�ƌ������CTCR �Ƒ��ݍ�p����@p.1577
- MHC �^���p�N��T �זE��K�ȕW�I�ɓ����悤�ɂ���@p.1579
- CD4 �����CD8 �⏕��e�̂�MHC �^���p�N�̕s�ϕ����Ɍ�������@p.1580
- �זE���Q��T �זE�́C�N���XI MHC �^���p�N�ƌ��������ȍזE���^���p�N�f�Ђɉ�������@p.1581
- �G���h�T�C�g�[�V�X�Ŏ�荞�܂�ăN���XII MHC �^���p�N�ƌ��������ȃ^���p�N�f�ЂɃw���p�[T �זE����������@p.1583
- �����𗧂�T �זE�͋��B���Ő��̑I������@p.1585
- ���ȃy�v�`�h�|MHC �����̂ɂ���Ċ�����������זE���Q��T �זE��w���p�[T �זE�̑啔���́C�����̉ߒ��ŋ��B���Ŕr�������@p.1586
- ���B�����ňُ����������鑟����ٓI�^���p�N������@p.1587
- MHC �^���p�N�̑��^�͂��̋@�\�Ő����ł���@p.1588
- �܂Ƃ߁@p.1588
���擪�s���E�I��\�ցA�͐߂̖ڎ��� �A�\�Q����
|
| 25��
�@�T�� |
25-5. �w���p�[T�זE�ƃ����p���̊������i10���ځj�@ p.1589
- ��������������זEd�͕����̋@�\��p����T �זE������������@p.1590
- T �זE�̊������͕��̃t�B�[�h�o�b�N�ɂ���Đ��䂳���@p.1591
- �K���Ɖu�����̐����̓G�t�F�N�^�[�w���p�[T �זE�̎�ނɂ���Č��܂�@p.1592
- TH1 �זE�͊��������}�N���t�@�[�W�����������C���ǔ������h������@p.1594
- B �זE��e�́iBCR�j�ւ̍R���̌�����B �זE�̊�������1 �i�K�ɂ����Ȃ��@p.1595
- �R�����ٓI�w���p�[T �זE�͂قƂ�ǂ�B �זE�̊������ɕs���ł���@p.1597
- ����̌n����B �זE��T �זE��ˑ��R�������ʂ���@p.1598
- �Ɖu�n�̎��ʕ��q�͋N���̌Â�Ig �X�[�p�[�t�@�~���[�ɑ����Ă���@p.1599
- �܂Ƃ߁@p.1600
- ���@���@p.1600
���擪�s���E�I��\�ցA�͐߂̖ڎ��� �A�\�Q����
|
| �� ���M���̍\���́u�S5��, �S25��, �S109��, �S1317����, �S1585�Łv���� |
| ���擪�s���E�I��\�ցA�͐߂̖ڎ��� �A�\�Q���� |
�I���̃��b�Z�[�W
���������ы��ނƂ��Ă̍זE�����ƍזE�̊w�сF�ȒP���L�ƃ��W�J���V���L���O��
�@�זE�̕��q�����w�́A��ؓ�ɂ͐i�܂Ȃ��E���t�������y�[�W���Ɛ������A�ҏW�Ɋւ�����p�ꌗ�̖����̐搶����ĂƂ������̂��Ȃƌ����Ă��܂����A������������t�Ƃ��Ă͉����Ɠs�����ǂ��B�ł��w���ɑ��āu������������Ăˁv�ƈʼn_�Ɍ����Ă��܂��������p���������B1000�ȏ���̍��ڂ����������ǂ�ł��Ă͓ǂݐi�߂�ɂ�ǂ��Ƃ͂����ƖY��Ă��܂��B�u�����č��鋳�t�̗���v�ł���B���{������Ƃ����X�͂ǂ̂悤�ɑΉ�����Ηǂ��̂��H�ƔY��ł��܂��B�����͂Ȃ�������̑Ή���̒�Ă͕K�v�ł��낤�B����ŁA�{�҂ł͍זE�����w�K�̈ꕔ�Ƃ��Ď�舵���Ă���B�m�����x���Ƃ͖��W�ɒN�������g�߂�����w�K�F����E�����E���̕K�v���ł���A���������Ɋ�Â��זE�����̎�舵���ł���B
�@�Ƃ���ŁA�זE�����L�b�g�� �u�v���ȕւ̋ɂ݁E�Ƃ������������s���Ă݂悤�E���茴���͌�ł����v�I�v �����ۂɉ\�Ƃ���B �_���؋��E����ǃ��W�b�N���K�v �u���A���e�B�[�Ɋ�Â��l����w�K�v�̎��H�ŁB ��I�Ȑ��������w�K�Ƃ��̓W�J���n���I�ɒ���B ���̍\���́A1)�T�v����A2)���Z�����A3)�����l�@�A4)���W�W�J�iWeb�x���w�K�E�o�[�`�����������ώ@�Ȃǁj����{�Ƃ��邪�A�A�����W�Ή�����ʓI�ł���B
�@���������Z�ł́A�w����ւ̔z���u���J�Ȏ��Z�w���v���ӎ�����邽�ߎ��ԓI����s�{�ӂȂ���l�@���ԁi�\�����ɂ��ώ@�]���j�Ȃǂ̒Z�k�����݉�����B�O�����ȋ��t�ւ̎x����A�{�����w�K�V�X�e���̌��ʓI�ȉ^�p�@�E�V���Ȋ���Ă��K�v�ł���B�܂�A�{�V�X�e���̖{���u���A���e�B�[�̊ȒP���L�Ɋ�Â��l����w�K�̎������v�ɂ͂���Ȃ�H�v���K�v�ƍl���Ă���B
�@�ȏ���ӂ݁A�܂��A���S�ȐV�K�u�Œ�t�v�̊J���Ɋ����Ɋ�Â��A�Ⴆ�A���w���x���ւ̍זE�����n�̓����A���̌o�܂��獂�Z�ł̕��S�y���Ƃ���������\�ł��낤�B�X�ɂ́A�זE�����L�b�g�ɂ��u���E���E��Ɉ�т������ފw�K�V�X�e���v�Ƃ������Ƃ������I�ɂ͗L���ł��낤�B���̎��ɂ͏�q�����u�זE�̓����F�זE���g�͉����Ă���H�v�ւ̑Ή������ՂȂ��̂ɂȂ��Ă��邱�Ƃ����҂������B�����̊ϓ_����A�V���ȁu���_�E�N�_�E�A�g�E���E���v�Ɋ�Â����̓��̒T�����K�v�Ȃ̂�������Ȃ��B
�@�Ō�Ɉꌾ�F���_�u�\���v�Ɋ�Â��זE�W�{�ώ@�@�ɂ͂Ƃ������N�����u�Ȃ�قǂˁI�v�ƌ����Ă����B�|�{�זE��������d�g�݁u�זE�|�{�R�v�f�v���������u���`�������I�v�A�זE�^�����݂�u�A���[�o�[�݂����I�v�A�זE�̊�{�I�Ȑ����ɂ́u���ꂾ���E�����Ɛ������āI�v�ł���B�C���t���ɑΖʂ��鋫�E�ʂ͍זE�V�[�g�Ɛ�������u������O�E����{���H�v�ƂȂ��Ă��܂��B����Ȃ���ȂŎ����w�K�͂Ƃ��������ՂɗL���ł���B�N�����D������ɂ��낢��Ȃ��Ƃ��l����E�b�������B������܂Ƃ߂�ƁuThe Cell�v�̂悤�ɂȂ�̂�������Ȃ��B
�@������O�Ȃ��Ƃ����������������Ă�͂胂�������A�ȒP���L�͍�����K�v�ۑ�ł���B���W�J���V���L���O�u�s�v�c���ˁE�Ȃ�قǂˁE�y�����ˁv���ĉ����Ă����w�K�̏���K�v�ł���B �w�K�҂Ƙb���������̓����l�������Ǝv���Ă���B�ꌾ�E�E���킹�ă~�R�g�A���t���u�_���؋��v�ł���B�����Ɍ����� Open Question ���Ă�����H�w�K�̏�̋��t�̎p���K�v�ł���B |